
Vorkommen des Soil-borne wheat mosaic virus (SBWMV-NY) in Schleswig-Holstein und dessen Bedeutung für den Weizenanbau
Occurrence of Soil-borne wheat mosaic virus (SBWMV-NY) in Schleswig-Holstein and its importance for wheat cultivation
Journal für Kulturpflanzen, 64 (12). S. 469–477, 2012, ISSN 1867-0911, DOI: 10.5073/JfK.2012.12.03, Verlag Eugen Ulmer KG, Stuttgart
Erhebungen zum Vorkommen bodenbürtiger Viren in Schleswig-Holstein zeigten, dass in diesem Bundesland das Soil-borne wheat mosaic virus (SBWMV) in Weizen- und Roggenanbaugebieten verbreitet ist. Während in Süddeutschland der Nebraska (N)-Stamm dieses Virus vorkommt, tritt in Norddeutschland der New York (NY)-Stamm auf. In den Infektionsherden der Getreidefelder werden verschiedene Winterweizensorten stark geschädigt. Um Anbauempfehlungen für die Kultivierung virusresistenten Weizens in Befallsregionen geben zu können, wurden ausgewählte Sorten unter Gewächshausbedingungen in infektiöser Erde eines betroffenen Standortes auf Resistenz getestet. Weiterhin erfolgte die Identifizierung des diagnostischen Markers Xgwm469-5D (153 bp- oder 155 bp-Allel) für das Sbm1-Resistenzgen gegen Furoviren im geprüften Weizenmaterial. Im Ergebnis dieser Untersuchungen können für den Weizenanbau in mit SBWMV-NY verseuchten Flächen die Sorten Farandole, Hyland, Hybery, Mirage und Carenius empfohlen werden.
Stichwörter: Bodenbürtige Furoviren, Soil-borne wheat mosaic virus-Stamm New York, Schleswig-Holstein, Winterweizen, Resistenz
The study of soil-borne viruses in Schleswig-Holstein shows an increasing distribution of Soil-borne wheat mosaic virus (SBWMV) in wheat and rye growing areas in this federal state of Germany. Whereas the Nebraska (N) strain of this virus was found in Southern Germany, the New York (NY) strain is present in Northern Germany. Selective wheat cultivars were screened for resistance to this virus under greenhouse conditions in an infested soil sample with the aim to develop recommendations for growers. Furthermore, the identification of the diagnostic marker Xgwm469-5D (153 bp- or 155 bp-allel) for the Sbm1-resistance gene against furoviruses was carried out in the wheat material under investigation. As a result of our work, the cultivation of the wheat cultivars Farandole, Hyland, Hybery, Mirage and Carenius is recommended for SBWMV-NY affected fields.
Key words: Soil-borne furoviruses, Soil-borne wheat mosaic virus-strain New York, Schleswig-Holstein, winter wheat, resistance
Das Soil-borne wheat mosaic virus (SBWMV) wurde 1919 in den USA entdeckt (McKinney, 1923) und erwies sich als hoch aggressives Pathogen in Weizen (Palmer und Brakke, 1975; Hunger et al., 1989). In Europa werden bodenbürtige Virosen an Weizen, Triticale und Roggen durch die beiden Furoviren Soil-borne cereal mosaic virus (SBCMV) und SBWMV sowie durch das Bymovirus Wheat spindle streak mosaic virus (WSSMV) verursacht. Für das SBWMV wurden entsprechend der geografischen Verbreitung unterschiedliche Stämme in den USA (Stamm ‚US-Nebraska–N‘, New York–NY‘, Oklahoma–OKL‘, Illinois–Il‘) und Isolate aus China, Japan und Europa (Deutschland und Polen) beschrieben (Shirako et al., 2000; Koenig und Huth, 2003; Fauquet et al., 2005; Trzmiel et al., 2012; Ziegler et al., 2012). Diese Viren werden durch Zoosporen des Protisten Polymyxa graminis Led. in die Pflanzenwurzeln übertragen und breiten sich im anfälligen Wintergetreide mit Beginn der Wachstumsphase im Frühjahr sehr schnell aus. Sie beeinträchtigen das Wachstum und die Bestockung der Pflanzen und führen zu beträchtlichen Ertragsverlusten im Wintergetreide (bis zu 88% im Weizen im Bundesstaat Kansas, USA), die durch Bodendesinfektion nicht nachhaltig zu hemmen sind (Eversmeyer, 1983). Eine erfolgreiche Bekämpfung bodenbürtiger Virosen kann deshalb nur durch den Anbau virusresistenter Sorten erzielt werden.
In Schleswig-Holstein wurden 2011 auf einer Fläche von 203 800 ha Weizen, auf 19 100 ha Roggen und auf 5800 ha Triticale angebaut (Beschreibende Sortenliste, 2012). In einigen Roggen- und Weizenanbaugebieten unterschiedlicher Regionen dieses Bundeslandes wurden seit 2006 wiederholt Furoviren nachgewiesen (Golecki et al., 2012). In diesen Befallsgebieten tritt der SBWMV-Stamm New York (NY) (Ziegler et al., 2012) mit hoher Aggressivität an Weizen und Roggen auf. Um Empfehlungen für die stabile Winterweizenproduktion in betroffenen Feldern geben zu können, wurde eine Auswahl von für diese Region empfohlenen Weizensorten auf Resistenz gegen das SBWMV-NY unter Gewächshausbedingungen getestet und ein Screening auf bekannte diagnostische Marker für Furovirusresistenz durchgeführt.
Basierend auf Beobachtungen von Landwirten und Pflanzenschutzberatern wurden seit 2006 in Getreideanbaugebieten unterschiedlicher Regionen Schleswig-Holsteins durch das Diagnoselabor der Landwirtschaftskammer Schleswig-Holstein in Kiel wiederholt bodenbürtige Viren in Weizen und Roggen diagnostiziert. Die Pflanzenproben aus verdächtigen Feldern wurden mittels DAS-ELISA mit polyklonalen Antiseren der Firma Loewe und der Deutschen Sammlung von Mikroorganismen und Zellkulturen GmbH (Leibniz-Institut DSMZ) und in der Reverse Transkriptase-Polymerase-Kettenreaktion (RT-PCR) geprüft. Auf Grund der aktuellen Befallssituation wurden Mitte Mai 2012 Pflanzen verschiedener Weizensorten von 6 Feldern aus der Umgebung der Dorfschaft Cashagen auf die bodenbürtigen Furoviren SBCMV und SBWMV untersucht.
Der serologische Nachweis auftretender Viren erfolgte mit DAS-ELISA und Julius Kühn-Institut (JKI)-eigenen polyklonalen Antiseren. Da beide Antiseren Kreuzreaktionen mit beiden Viren zeigen, wurde für die sichere Differenzierung des SBWMV der spezifische monoklonale Antikörper (mAK) 4G4 (Isotyp IgG3, ƙ) im DAS-ELISA eingesetzt (Rabenstein et al., 2005).
Der elektronenmikroskopische Nachweis von Viruspartikel des SBWMV-NY im Blattsaft infizierter Feldpflanzen der Sorte Primus erfolgte sowohl mittels Immunelektronenmikroskopie (ISEM-Technik) durch Dekoration der Partikel unter Einsatz des gereinigten mAK 4G4 als auch mittels Immunogold-Markierungen unter Verwendung des mAK 4G4. Die Präparate wurden jeweils mit 2% Uranylacetat kontrastiert. Der molekularbiologische Nachweis der Furoviren fand durch RT-PCR wie folgt statt: Gesamt-RNA wurde unter Nutzung des Minikits Bio&SELL RNA Tri-Flüssig isoliert und die cDNA synthetisiert. Die RT-PCR verlief nach einem Standardprotokoll (3 min bei 96°C, 35 Zyklen mit 30 s bei 96°C, 75 s 56°C, 60 s bei 72°C und mit einer abschließenden Extension von 10 min bei 72°C) unter Verwendung virusspezifischer Primer für das SBWMV (PGRV4a-5’CTGCGACTCACGCTTACATA3‘, PGRV
4b-5’TAACCGCTTTGGGATGATAG3‘) mit einem PCR-Produkt von 680 bp und für das SBCMV (SBCMVfor_RNA2-
5’ACTTACCCATTTAGGTGTAA3‘, SBCMVrev_RNA2-5’TTATAATCACGCAAGTACCT3‘) mit einem PCR-Produkt von 978 bp (Fomitcheva et al., 2008).
Die Erhebung von Resistenzreaktionen gegen das SBWMV-NY wurde an ausgewählten Weizensorten unterschiedlicher Züchterfirmen, an Weizen-Linien und an der Gersten-Landrasse Muju covered 2 aus Korea, die Resistenz gegen die Gerstengelbmosaikviren besitzt (Ordon, 1999), für das SBWMV jedoch anfällig ist (Kastirr et al., 2008), vorgenommen (Tab. 1). Die Resistenzprüfung erfolgte unter kontrollierten Gewächshausbedingungen (Temperaturen zwischen 17°C bis 23°C, Luftfeuchtigkeit ca. 50% und umweltabhängiger Zusatzbeleuchtung bis zu 8 Stunden bei 10 klux). Je Prüfglied wurden 25 Körner in Multitopfplatten mit SBWMV-NY kontaminierter Erde vom Schlag Bormkoppel aus der Region Cashagen ausgelegt. Der Virusnachweis wurde mittels DAS-ELISA drei, fünf und acht Wochen nach Aussaat (wpi) in Wurzeln, Spross und Blättern von 5 Einzelpflanzen je Sorte durchgeführt, wofür Pflanzensaft aus den Wurzeln (bei großen Wurzelballen Querschnittsprobe von ca. 2 cm Länge), aus dem unteren Sprossteil (ca. 2 cm) und dem oberen Sprossteil mit Blattansatz (ca. 2 cm) in der Walzenpresse mit 400 µl Extraktionspuffer gewonnen wurde. Nach 10 Wochen wurden weiterhin die Blätter von 10 Einzelpflanzen je Prüfglied auf Virusbefall getestet.
Tab. 1. Material für die Resistenzprüfung gegen SBWMV-NY
Kultur | Sorte/Linie | Züchter |
Winterweizen | Akteur | Deutsche Saatveredelung AG |
Potenzial | Deutsche Saatveredelung AG | |
Carenius | Dieckmann GmbH&Co.KG | |
Farandole | Dr. Peter Franck Pflanzenzucht Oberlimpurg | |
Plutos | Dr. Hermann Strube | |
Julius VSS | KWS LOCHOW GMBH | |
Ritmo | Limagrain-Nederland B.V. | |
Tuareg | NORDSAAT Saatzuchtgesellschaft | |
Hyland | NORDSAAT Saatzuchtgesellschaft | |
Hybery | SAATEN UNION RECHERCHE SAS | |
Meister | RAGT 2N | |
Mirage | RAGT 2N | |
JB Asano | Saatzucht Josef Breun GmbH&Co.KG | |
Event | Saatzucht Josef Breun GmbH&Co.KG | |
Inspiration | Saatzucht Josef Breun GmbH&Co.KG | |
Tobak | W. von Borries-Eckendorf GmbH&Co.KG | |
Rene Leblond | Frankreich (PI 352294) | |
WW Gi 03–27 | NORDSAAT Saatzuchtgesellschaft | |
(F1 Tremie × Aztek) | (Frankreich INRA) | |
Cltr 17886 | USA, South Dakota | |
(PI15092 × T. speltoides) | ||
Gerste | Muju covered 2 (sechszeilig) | Landrasse aus Korea (Ordon, 1999) |
Außerdem erfolgte das Screening genomischer DNA des geprüften Weizenmaterials zur Identifizierung diagnostischer Marker (153 bp- oder 155 bp-Allele) im Bereich des Sbm1-Gens mit dem Mikrosatellitenmarker Xgwm469-5D. Die genomische DNA wurde mittels CTAB-Methode (Graner et al., 1991) isoliert. Eine zusätzliche Extraktion mit Chloroform:Isoamylalkohol (24:1) wurde vor der Ethanolfällung vorgenommen. Die PCR wurde in einem AB9700 Thermal Cycler (Applied Biosystems) mit Fluoreszenz-markiertem Cy5 Vorwärts- und unmarkiertem Rückwärts-Mikrosatellitenprimern durchgeführt. Die Analyse der PCR-Produkte erfolgte mit dem CEQTM 8000 Genetic Analysis System (Perovic et al., 2009a).
In Schleswig-Holstein werden seit 2006 in unterschiedlichen Getreideanbaugebieten bodenbürtige Viren in Winterweizen und Winterroggen nachgewiesen (Abb. 1). In den Gemeinden Wittenbergen im Kreis Steinburg und Woltersdorf im Kreis Herzogtum Lauenburg wurden 2006 Befallsherde in Roggenfeldern beobachtet. Im Norden des Bundeslandes an der Grenze zu Dänemark wurden 2009 Virus kontaminierte Roggenfelder in der Gemeinde Weesby des Kreises Schleswig-Flensburg sowie infizierte Weizenschläge im Hauke-Haien-Koog Nordfrieslands an der Westküste Schleswig-Holsteins erfasst. In Umgebung der südöstlichen Dorfschaft Cashagen im Kreis Ostholstein wurden 2009 und wiederholt 2012 bodenbürtige Weizenvirosen registriert.

Abb. 1. Verbreitung des Soil-borne wheat mosaic virus Stamm New York (SBWMV-NY) in Schleswig-Holstein (Image © 2012 Geo Content, Google Earth).
Der Virusbefall in den infizierten Weizenpflanzen wurde mittels serologischer und molekularbiologischer Nachweistechniken differenziert. Im Resultat dieser Analysen wurde nachgewiesen, dass in den untersuchten Flächen das Furovirus SBWMV in Winterweizen und Winterroggen dominiert (Tab. 2).
Tab. 2. Nachweis des SBWMV in Winterroggen und Winterweizen mittels RT-PCR in unterschiedlichen Standorten Schleswig-Holsteins in den Jahren 2006 bis 2012
Jahr | Standort | Kultur | Virusnachweis |
2006 | Wittenbergen | Roggen | + |
Woltersdorf | Roggen | + | |
2009 | Hauke-Haien-Koog | Weizen | + |
Weesby | Roggen | + | |
Cashagen | Weizen | + | |
2012 | Cashagen | Weizen | + |
Die Identifizierung des SBWMV erfolgte weiterhin durch spezifische Markierung der Viruspartikel aus infizierten Blättern von Feldpflanzen der Sorte Primus. Die elektronenmikroskopische Darstellung der stäbchenförmigen Partikel mit Zentralkanal zeigte die Existenz von zwei Partikellängen. In den Blattsaftpräparaten wurden Virionen im Längenbereich von 125 nm bis 175 nm und von 200 nm bis 340 nm beobachtet (Abb. 2A). Zum Nachweis der Identität der Partikel mit dem SBWMV wurde eine Partikeldekoration mit dem spezifischen mAK 4G4 vorgenommen (Abb. 2B) und eine Immunogold-Markierung durchgeführt (Abb. 2C).
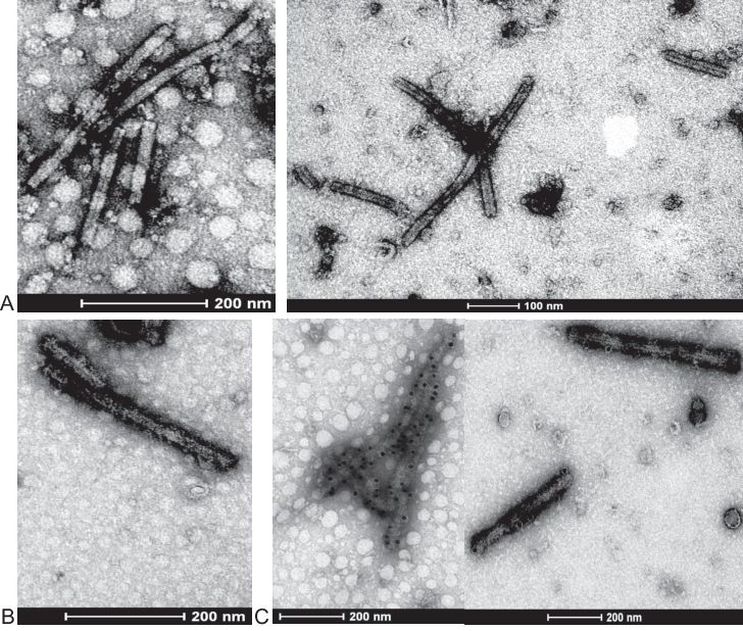
Abb. 2. Elektronenmikroskopische Darstellung von stäbchenförmigen Partikeln mit Zentralkanal des SBWMV-NY aus infizierten Blättern von Feldpflanzen der Weizensorte Primus A – undekorierte Partikel in zwei Längenbereichen, B – mAK 4G4 dekorierte SBWMV Partikel, C – Immunogold-Markierung mit mAK 4G4.
Im Ergebniss dieser Untersuchungen wurden eindeutig spezifisch markierte Viruspartikel des SBWMV identifiziert.
Diese Befunde wiesen für Deutschland eine zweite Region der Verbreitung dieses Furovirus aus. Im Jahr 2002 wurde in Baden-Württemberg am Standort Heddesheim das erste Mal ein Furovirus mit hoher Aggressivität an Winterweizen beobachtet (Koenig und Huth, 2003), welches als Nebraska-Stamm identifiziert wurde.
Die SBWMV-Isolate aus den Befallsregionen von Schleswig-Holstein wurden vergleichenden Sequenzanalysen unterzogen und konnten dem NY-Stamm des SBWMV zugeordnet werden (Ziegler et al., 2012).
Mitte Mai 2012 wurden sechs Weizenschläge in der Region um Cashagen (Tab. 3) besichtigt, um den Umfangs des Virusbefalls und dessen Einfluss auf den Weizenanbau in dieser Region bewerten zu können. Wie aus Tab. 3 ersichtlich ist, etablierten sich die Pathogenpopulationen aus pilzlichem Vektor (P. graminis) und dem SBWMV-NY sowohl in den nährstoffreichen Mineralböden mit hoher organischer Humusmasse (Mutterboden, anmooriger Boden) als auch in den leichteren Mineralböden (Schwemmboden, sandiger Lehm).
Tab. 3. Nachweis der SBWMV-NY-Infektion von Winterweizen mittels DAS-ELISA in unterschiedlichen Böden der südöstlichen Befallstandorte Schleswig-Holsteins
Schlag | WW-Sorte | Bodentyp | Virustiter (E405nm) | PCR | ||
SBCMV | SBWMV | SBWMV | ||||
1 | Bormkoppel | Primus | sandiger Lehm | 0,0 | 1,2 | + |
2 | Goldher-Hehlsen I | Farandole | sandiger Lehm | 0,0 | 0,0 | – |
3 | Langkoppel | Kredo | Mutterboden | 0,0 | 1,1 | + |
4 | Kredo | Schwemmboden | 0,0 | 1,0 | + | |
5 | Hundekamp | Diskus | anmooriger Boden | 0,0 | 1,1 | + |
6 | Heidkoppel | Kredo | sandiger Lehm | 0,0 | 1,2 | + |
Das SBWMV-NY erreichte in den Feldpflanzen der anfälligen Sorten hohe Virustiter. Nur am Standort Goldher-Hehlsen I, wo die Sorte Farandole angebaut wurde, konnte in diesem Jahr kein Virusbefall nachgewiesen werden. RT-PCR-Analysen bestätigten diese Ergebnisse (Abb. 3). Das SBCMV wurde in den Pflanzen dieser viruskontaminierten Felder nicht nachgewiesen.
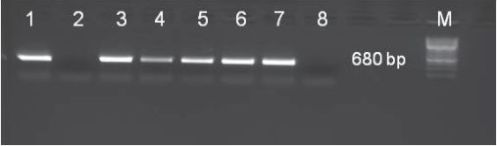
Abb. 3. RT-PCR-Nachweis des SBWMV-NY (Fragmentgröße 680 bp) in verschiedenen Winterweizensorten unterschiedlicher Weizenschläge in Umgebung von Cashagen im Jahr 2012 (1–6: Weizenschläge (s. Tab. 3), 7: positive Viruskontrolle, 8: negative PCR-Kontrolle, M: Molekülmassemarker (GeneRuler, 1 kb, Fermentas).
In vier der besichtigten Felder existierten entlang der Bearbeitungsrichtung bereits größere Infektionsherde (Abb. 4A). In diesen Schlägen zeigten die Weizensorten Primus, Kredo und Diskus starke Virussymptome. Im Vergleich zu den symptomlosen Pflanzen war die Wuchshöhe infizierter Pflanzen Mitte Mai bereits um ca. 50% reduziert (Abb. 4B). Virus befallene Pflanzen hatten kaum noch symptomfreie Blätter (Abb. 4C) und oft nur einen oder zwei Bestockungstriebe (Abb. 4D).

Abb. 4. Infektion der Sorte Primus durch SBWMV-NY im Winterweizenschlag Bormkoppel A – Infektionsherd im Feld, B – Wuchsdepressionen infizierter Pflanzen im Vergleich zu gesunden Pflanzen, C – Strichel-Mosaiksymptome an Primus, D – geringe Bestockung.
Unsere Beobachtungen weisen darauf hin, dass die Weizenproduktion in diesen Regionen vom Anbau SBWMV-resistenter Sorten profitieren kann.
Das SBWMV-NY infiziert eine Reihe zugelassener Weizensorten (Tab. 3). Um Anbauempfehlungen für die Befallsgebiete geben zu können, wurden Sorten mit Resistenz gegen Furoviren (Farandole, Mirage, Carenius), anfällige Sorten (JB Asano, Meister, Event) sowie die gegen das SBWMV anfällige Gersten-Landrasse Muju covered 2 in infektiöser Erde des Schlages Bormkoppel kultiviert. Die Virusinfektion wurde mittels DAS-ELISA in Wurzeln, Spross und Blättern 3, 5 und 8 wpi erfasst. Zehn Wochen nach Aussaat wurde die Virusinfektion in den Blättern weiterer Pflanzen abschließend geprüft. Es wurde ersichtlich (Abb. 5), dass in den Wurzeln der anfälligen Weizensorten die Virusinfektion bereits 3 wpi nachweisbar war und in den Sorten JB Asano und Event zu diesem Zeitpunkt bereits den Spross erfasst hatte. Zwei Wochen später enthielten Wurzeln, Spross und Blätter dieser Sorten hohe Virustiter, die sich auch 8 wpi bestätigten. Zehn Wochen nach Aussaat waren die Blätter der Einzelpflanzen (n = 10) von JB Asanao zu 100%, von Event zu 92% und von Meister zu 83% infiziert. Auch die Furovirus anfällige sechszeilige Gersten-Landrasse (G) Muju covered 2 zeigte hohe Virustiter in allen Pflanzenteilen. In den drei resistenten Sorten wurde das SBWMV-NY in den Wurzeln erst 3 bis 5 wpi nachgewiesen. Die Virusausbreitung in Spross und Blätter war im Vergleich zu den anfälligen Sorten unter optimalen Gewächshausbedingungen stark gehemmt. Geringe Virustiter im Spross wurden nur vereinzelt 8 wpi beobachtet.
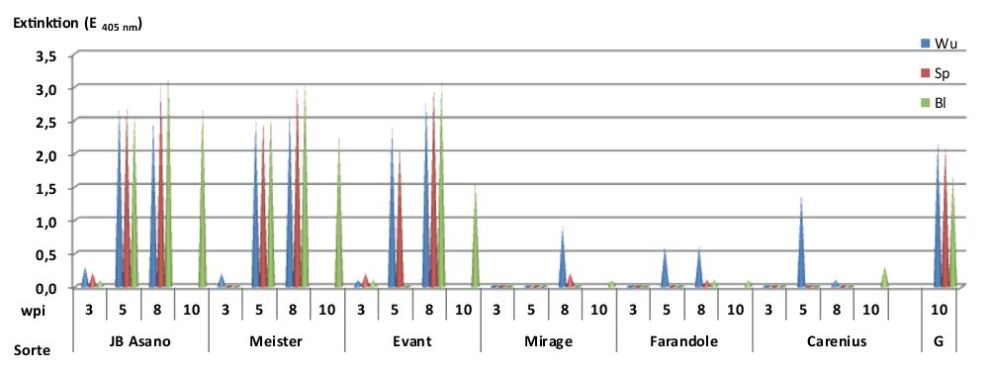
Abb. 5. Vergleich der Resistenzreaktionen von anfälligen und resistenten Weizensorten und der anfälligen Gersten-Landrasse (G) gegen das SBWMV-NY nach Kultivierung in infektiöser Erde des Schlages Bormkoppel unter Gewächshausbedingungen im Zeitraum von 3 bis 10 wpi, Wu – Wurzel-, Sp – Spross-, Bl – Blattinfektion.
Weitere Weizensorten und -linien zeigten eine deutliche Resistenzdifferenzierung (Abb. 6). Die Linie Cltr17886, die Sorten Rene Leblond, Hyland, Hybery und Plutos und die DH-Linie WW Gi 03-27 wiesen bis 8 wpi nur geringe Virustiter in den Wurzeln auf. In der darauf folgenden Inkubationszeit erfolgte keine Virusausbreitung in Spross und Blätter. In diesen Weizenformen liegt offensichtlich eine Translokations(Ausbreitungs)-Resistenz gegen das Virus vor, deren genetischer Hintergrund noch unbekannt ist (Hariri et al., 1987; Myers et al., 1993; Huth et al., 2007; Lyons et al., 2008). In den anfälligen Weizensorten hingegen wurde ab 5 wpi Virusbefall mit mittleren und hohen Virustitern auch in Spross und Blättern nachgewiesen.
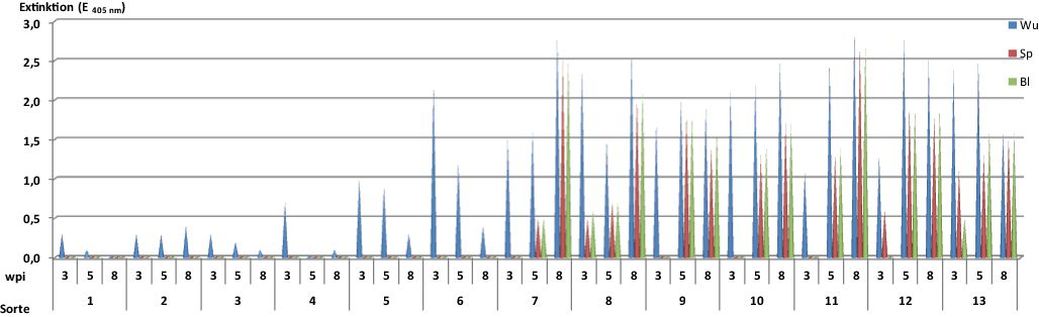
Abb. 6. Prüfung der Resistenz von zugelassenen Weizensorten und Resistenzträgern gegen das SBWMV-NY in infektiöser Erde unter Gewächshausbedingungen im Zeitraum von 3 bis 8 Wochen nach Aussaat (wpi) Wu – Wurzel-, Sp – Spross-, Bl – Blattinfektion; 1 Cltr 17886; 2 Rene Leblond; 3 Plutos; 4 WW Gi 03-27; 5 Hyland; 6 Hybery; 7 Akteur; 8 Ritmo; 9 Tuareg; 10 Inspiration; 11 Potenzial; 12 Tobak; 13 Julius VSS.
Das Material wurde einer Analyse zur Identifizierung des Resistenzgen-Locus Sbm1, der mit der Furovirusresistenz in Weizen korreliert (Kanyuka et al., 2004; Bass et al., 2006; Perovic et al., 2009b), unterzogen. Dieses Resistenzgen wurde auf dem Chromosom 5 des D-Genoms im hexaploiden Weizen kartiert und wird monogen vererbt. Es kann durch den Mikrosatellitenmarker Xgwm469-5D nachgewiesen werden. In resistenten Genotypen weist der Xgwm469-5D-Marker ein 153 bp- oder ein 155 bp-Allel nach und in anfälligen Weizen ein Null-Allel in diesen Positionen.
In der Tab. 4 wurden die Ergebnisse zur Identifizierung diagnostischer Marker den Extinktionswerten (E405 nm) nach achtwöchiger Inkubationszeit gegenüber gestellt.
Tab. 4. Screening von Weizensorten 8 wpi auf die 153 bp- oder 155 bp-Allele für Resistenz gegen Furoviren mit dem diagnostischen Marker Xgwm469-5DL (– Nullallel bei anfälligen Sorten, A-anfällig, R-resistent)
Nr. | Akzession/Sorte | SSR-Analyse | DAS-ELISA (8 wpi) (E405 nm) | ||||
Xgwm469-5DL-Allele | erwarteter | Wurzel | Spross | Blatt | |||
Abbildung 5 | |||||||
1 | JB Asano | – | A | 2,5 | 3,0 | 3,2 | |
2 | Meister | – | A | 2,6 | 3,0 | 3,1 | |
3 | Event | – | A | 2,8 | 3,0 | 3,1 | |
4 | Mirage | 153 | R | 0,9 | 0,2 | 0,0 | |
5 | Farandole | 155 | R | 0,6 | 0,1 | 0,1 | |
6 | Carenius | 155 | R | 0,1 | 0,0 | 0,0 | |
Abbildung 6 | – | ||||||
1 | Cltr 17886 | 153 | R | 0,1 | 0,0 | 0,0 | |
2 | Rene Leblond | 155 | R | 0,4 | 0,0 | 0,0 | |
3 | Plutos | 155 | R | 0,1 | 0,0 | 0,0 | |
4 | WW Gi 03–27 | 153 | R | 0,1 | 0,0 | 0,0 | |
5 | Hyland | 155 | R | 0,3 | 0,0 | 0,0 | |
6 | Hybery | 155 | R | 0,4 | 0,0 | 0,0 | |
7 | Akteur | – | A | 2,8 | 2,5 | 2,5 | |
8 | Ritmo | – | A | 2,6 | 2,0 | 2,1 | |
9 | Tuareg | – | A | 1,9 | 1,4 | 1,6 | |
10 | Inspiration | – | A | 2,5 | 1,7 | 1,7 | |
11 | Potenzial | – | A | 2,9 | 2,7 | 2,7 | |
12 | Tobak | – | A | 2,5 | 1,8 | 1,9 | |
13 | Julius VSS | – | A | 1,6 | 1,5 | 1,6 | |
Die Resistenzreaktionen dieser Weizensorten stimmten mit den Ergebnissen des Nachweises der diagnostischen Marker sehr gut überein.
Die Resistenzprüfungen in den zugelassenen Weizensorten zeigten, dass resistente Sorten (Farandole, Hyland, Hybery, Mirage und Carenius) vorhanden sind, welche in Befallsflächen Schleswig-Holsteins die Weizenproduktion sichern können. Andere Resistenzquellen sind Linienmaterial und die in der Zulassung befindliche Sorte Plutos.
Die Gefahr für den Weizenanbau durch das Auftreten bodenbürtiger Virosen wurde bisher in Deutschland als gering eingeschätzt. Mit dem Auftreten des SBWMV-NY in Schleswig-Holstein relativiert sich diese Einschätzung.
Unsere Beobachtungen und neueste Berichte aus Polen (Trzmiel et al., 2012) belegen, dass das SBWMV in Europa weiter verbreitet ist, als bisher bekannt war. Die polnischen Autoren wiesen in der Region Westpolens (südliches Wielkopolska) in Triticalepflanzen das SBWMV-Nebraska nach (Abb. 7).
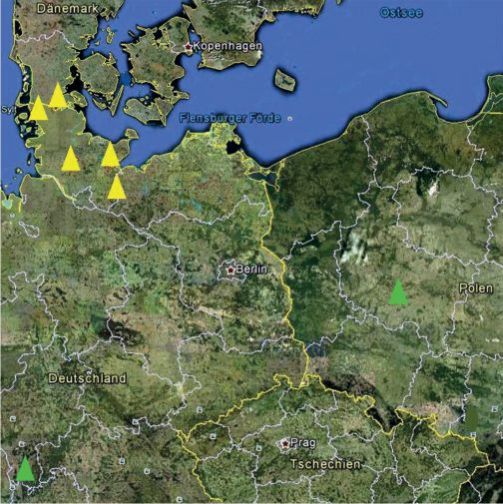
Abb. 7. Verbreitung des Nebraska-(grüne Dreiecke) und des New York-Stammes (gelbe Dreiecke) des SBWMV in Europa (Image © 2012 Geo Content, Google Earth).
Um epidemiologische Studien zur Verbreitung unterschiedlicher Stämme des SBWMV durchführen zu können, sollte das Vorkommen dieses Virus in Europa detailliert untersucht werden.
Die Resistenzzüchtung gegen Furoviren ist für die verschiedenen Getreidearten unterschiedlich entwickelt. Für den Weizen als ökonomisch bedeutsamste Art wurden weltweit Resistenzquellen gegen bodenbürtige Viren evaluiert (Modawi et al., 1982; Merkle und Smith, 1983; Cox et al., 1994; Barbosa et al., 2001; Bayles und Napier, 2002), Resistenzloci in den D- und B-Genomen kartiert und erste diagnostische Marker für Allele, die mit der Virusresistenz korrelieren, entwickelt (Bass et al., 2006; Perovic et al., 2009b; Maccaferri et al., 2011). Es wurden auf dem langen Arm von Chromosom 5 des D-Genoms und auf dem kurzen Arm des Chromosom 2 des B-Genoms Resistenzgene lokalisiert, die durch spezifische diagnostische Marker im Zuchtmaterial von Weich- (Triticum aestivum) und Hartweizen (Triticum durum) identifiziert werden können. Wie unsere Untersuchungen zur Sortenresistenz gegen das SBWMV-NY zeigen, gibt es auch in Deutschland zugelassene Winterweizensorten (Farandole, Hyland, Hybery, Mirage, Carenius) mit Resistenz gegen Furoviren.
Die in Europa im Anbau befindlichen Roggen- und Triticalesorten sind jedoch sehr anfällig für Furoviren (Kastirr et al., 2002, 2005). Im Anatolischen Genpool des Roggens konnten Resistenzquellen gegen bodenbürtige Viren nachgewiesen werden. Nach Individualauslese aus Pärchenkreuzungen erfolgte die Selektion von Vollgeschwisterfamilien mit erhöhtem Resistenzniveau. Einige Resistenz-Donoren wurden in Elite-Zuchtmaterial eingekreuzt und dienen der Entwicklung virusresistenter Roggensorten (Kastirr et al., 2011).
In den bisher untersuchten europäischen Triticalesorten konnten keine Resistenzen gegen Furoviren gefunden werden.
Aus der Anfälligkeit einiger Gerstenformen für SBWMV-N/-NY leitet sich eine mögliche Beeinträchtigung des Gerstenanbaus durch das SBWMV ab.
Aus dem Auftreten des SBWMV-NY in Schleswig-Holstein und des SBWMV-N in Südpolen ergibt sich die Notwendigkeit des Monitorings bodenbürtiger Viren in Europa zur Identifizierung neuauftretender Virusstämme und zur Aufklärung ihrer Bedeutung für den Getreideanbau. Vor der Züchtungsforschung steht die Aufgabe, Resistenzquellen in Roggen und Triticale zu evaluieren und in die Sortenentwicklung zu integrieren, das Spektrum resistenter Weizensorten zu erweitern und die Bedeutung des SBWMV für die Kulturgerste zu untersuchen.
Wir danken der Fachagentur Nachwachsende Rohstoffe e.V. (FNR) beim Bundesministerium für Ernährung, Landwirtschaft und Verbraucherschutz (BMELV) für die Förderung des Projektes FKZ: 22019311 im Rahmen des Programmes „Nachwachsende Rohstoffe“. Unser Dank gilt Martje Katarina Ratzow (Landwirtschaftliche Unternehmensberatung Jan Wagner) für die Organisation der Probenahme in den Befallsstandorten Schleswig-Holsteins. Wir danken den in Tab. 1 aufgeführten Züchtern für die Bereitstellung des Saatgutes. Für die Bereitstellung von spezifischen Antiseren und monoklonalen Antikörpern danken wir Herrn Dr. Frank Rabenstein. Für die elektronenmikroskopischen Untersuchungen sagen wir Frau Elke Zimmermann, für die Identifizierung der diagnostischen Marker Frau Marlis Weilepp und für die Befallserfassung Frau Viola Papke Dank.
Barbosa, M.M., L. R. Goulart, A. M. Prestes, F.C. Juliatti, 2001: Genetic control of resistance to soil-borne wheat mosaic virus in Brazilian cultivars of Triticum aestivum L. Thell. Euphytica 122, 417-422.
Bass, C., R. Hendley, M.J. Adams, K.E. Hammond-Kosack, K. Kanyuka, 2006: The Sbm1 locus conferring resistance to Soil-borne cereal mosaic virus maps to a gene-rich region on 5DL in wheat. Genome 49, 1140-1148.
Bayles, R.A., B. Napier, 2002: Tolerance of wheat varieties to Soil-borne wheat mosaic virus (SBWMV). London, UK, HGCA Project Report No. 278, Home-Grown Cereals Authority.
Bundessortenamt, 2012: Beschreibende Sortenliste Getreide, Mais, Öl- und Faserpflanzen, Leguminosen, Rüben, Zwischenfrüchte. ISSN 2190-6130, 143.
Cox, T.S., M.E. Sorrells, G.C. Bergstrom, R.G. Sears, B.S. Gill, E.J. Walsh, S. Leath, J.P. Murphy, 1994: Registration of KS92WGRC21 and KS92WGRC22 hard red winter wheat germplasms resistant to wheat streak mosaic virus and powdery mildew. Crop Sci. 34, 546.
Eversmeyer, M.G., 1983: Effect of soil fumigation on occurrence and damage caused by Soilborne wheat mosaic. Plant Disease 67 (9), 1000-1002.
Fauquet, C.M., M.A. Mayo, J. Maniloff, U. Desselberger, L.A. Ball, (Eds.) 2005: Furovirus. In: ‚The eighth report of the international committee on taxonomy of viruses‘, 1027-1032.
Fomitcheva, V., U. Kastirr, A. Habekuss, T. Kühne, 2008: Diagnostic multiplex RT -PCR analysis for the detection of soil-borne mosaic viruses and their natural vector Polymyxa graminis. Quedlinburg, Germany, Proceedings of the Seventh Symposium of the International Working Group on Plant Viruses with Fungal Vectors (IWGPVFV), ed. Rush, C.M., 43-48.
Golecki, B., M. Wunderlich, S. Opitz, 2012: Vorkommen und Nachweis bodenbürtiger Viren an Roggen und Weizen im Bundesland Schleswig-Holstein. 44. Tagung des DPG-Arbeitskreises „Viruskrankheiten der Pflanzen“, 08.–09. März 2012, Quedlinburg, Julius Kühn-Institut, Vortrag.
Graner, A., A. Jahoor, J. Schondelmaier, H. Siedler, K. Pillen, G. Fischbeck, G. Wenzel, R.G. Herrmann, 1991: Construction of an RFLP map in barley. Theor. Appl. Genet. 83, 250-256.
Hariri, D., M. Courtillot, P. Zaoui, H. Lapierre, 1987: Multiplication of soilborne wheat mosaic virus (SBWMV) in wheat roots infected by a soil carrying SBWMV and wheat yellow mosaic virus (WYMV), Agronomie 7 (10), 789-796.
Hunger, R.M., C.R. Armitage, J.L. Sherwood, 1989: Effects of wheat soilborne mosaic virus on hard red winter wheat. Plant Dis. 73, 949-952.
Huth W., R. Götz, D.-E. Lesemann, 2007: Unterschiedliche Formen der Resistenz gegen bodenbürtige Viren des Weizens. Ges. Pflanzen 59, 29-39.
Kanyuka, K., D. Lovell, O.P. Mitrofanova, K. Hammond-Kosack, M.J. Adams, 2004: A controlled environment test for resistance to Soil-borne cereal mosaic virus and its use to determine the mode of inheritance of the resistance in the UK wheat variety Cadenza, and to screen diverse Triticum monococcum genotypes for potential sources of improved disease resistance. Plant Pathol. 53, 154-160.
Kastirr, U., F. Rabenstein, T. Kühne, 2002: Investigation of epidemiology of Soil-borne cereal mosaic virus in the German country Saxony-Anhalt. Proceedings of the Fifth Symposium of the International Working Group on Plant Viruses with Fungal Vectors, eds. Rush, C.M., U. Merz 2002, 119-122.
Kastirr, U., F. Rabenstein, T. Kühne, 2005: Epidemiological aspects of soil-borne viruses of wheat, triticale and rye in Germany. Bologna, Italy, Proceedings of the Sixth Symposium of the International Working Group on Plant Viruses with Fungal Vectors, ed. Rush, C.M, 2005, 132-136.
Kastirr, U., F. Ehrig, V.W. Fomitcheva, A. Habekuss, T. Kühne, 2008: Investigation of furovirus virulence in winter barley. Quedlinburg, Germany, Proceedings of the Seventh Symposium of the International Working Group on Plant Viruses with Fungal Vectors (IWGPVFV), ed. Rush, C.M, 2008, 117-121.
Kastirr, U., E. Bauer, B. Schmiedchen, C. Pietsch, V. Korzun, P. Wilde, 2011: Resistenz gegen die bodenbürtigen Viren Soil-borne cereal mosaic virus (SBCMV) und Wheat spindle streak mosaic virus (WSSMV) bei Winterroggen. Tagung der AG Krankheitsbekämpfung und Resistenzzüchtung in Getreide, Hülsenfrüchten und Raps, Fulda, GPZ-Homepage, http://www.gpz-online.de/ag/07.html.
Koenig, R., W. Huth, 2003: Natural infection of wheat by the type strain of Soil-borne wheat mosaic virus in a field in Southern Germany. Europe Journal of Plant Pathology 109, 191-193.
Lyons, R., K.E. Hammnd-Kosack, K. Kanyuka, 2008: Identification and characterization of a novel efficient resistance response to the furoviruses SBWMV and SBCMV in barley. Molecular Plant-Microbe Interactions 21, 1193-1204.
Maccaferri, M., C. Ratti, C. Rubies-Autonell, V. Vallega, A. Demontis, S. Stefanelli, R. Tuberosa, M.C. Sanguineti, 2011: Resistance to Soil-borne cereal mosaic virus in durum wheat is controlled by a major QTL on chromosom arm 2BS and minor loci. Theor. Appl. Genet. 123, 527-544.
Merkle, O.G., E.L. Smith, 1983: Inheritance of resistance to soil-borne mosaic in wheat. Crop Sci. 23, 1075-1076.
Modawi, R.S., E.G. Heyne, D. Brunetta, W.G. Willis, 1982: Genetic studies of field26 reaction to wheat soil-borne mosaic virus. Plant Dis. 66, 1183-1184.
Myers, L.D., J.L. Sherwood, W.C. Siegerist, R.M. Hunger, 1993: Temperature influenced virus movement in expression of resistance to Soilborne wheat mosaic virus in hard red winter wheat (Triticum aestivum). Phytopathology 83, 548-551.
McKinney, H.H., 1923: Investigation of the rosette disease of wheat and its control. J. Agr. Res. 23, 771-800.
Ordon, F., 1999: Markergestützte Selektion in der Resistenzzüchtung beim Getreide – unter besonderer Berücksichtigung des Pathosystems Gerste (Hordeum vulgare L.) – Bymoviren (BaMMV, BaYMV, BaYMV-2). Aachen, Shaker Verlag, Berichte aus der Agrarwissenschaft, 131 S.
Palmer, L.T., M.K. Brakke, 1975: Yield reduction in winter wheat infected with soilborne wheat mosaic virus. Plant Dis. Rep. 59, 469-471.
Perovic, P., K. Kanyuka, F. Ordon, 2009a: Disease resistance. Soil-borne cereal mosaic virus (SBCMV) resistance in bread wheat. http://maswheat.ucdavis.edu/protocols/sbcmv/.
Perovic, D., J. Förster, P. Devaux, D. Hariri, M. Guilleroux, K. Kanyuka, R. Lyons, J. Weyen, D. Feuerhelm, U. Kastirr, P. Sourdille, M. Röder, F. Ordon, 2009b: Mapping and diagnostic marker development for soil-borne cereal mosaic virus resistance in bread wheat. Molecular Breeding 23 (4), 641-653, ISSN/ISBN 1380-3743.
Rabenstein, F., H. Mühlheim, U. Kastirr, T. Kühne, 2005: Monoclonal antibodies for differentiation between Soil-borne cereal mosaic virus and Soil-borne wheat mosaic virus. Bologna, Italy, Proceedings of the Sixth Symposium of the International Working Group on Plant Viruses with Fungal Vectors, ed. Rush, C.M., 2005, 53-56.
Shirako, Y., N. Suzuki, R.C. French, 2000: Similarity and divergence among viruses in the genus Furovirus. Virology 270, 201-207.
Trzmiel, K., M. Jezewska, A. Zarzynska, 2012: First report of Soil-borne wheat mosaic virus (SBWMV)-infecting triticale in Poland. J. Phytopathol. DOI: 10.1111/j.1439-0434.2012.01952.x.
Ziegler, A., B. Golecki, U. Kastirr, 2012: Occurrence of the New York strain of Soil-borne wheat mosaic virus in Northern Germany. J. Phytopathol., (early view), DOI: 10.111jph.12050.

 Suchen
Suchen