
Genetic diversity among North African faba bean landraces for competitive ability against weeds
Genetische Diversität Nordafrikanischer Ackerbohnen-Landrassen für Konkurrenzkraft gegen Unkraut
Journal für Kulturpflanzen, 70 (5). S. 145–157, 2018, ISSN 1867-0911, DOI: 10.1399/JfK.2018.05.01, Verlag Eugen Ulmer KG, Stuttgart
The genetic diversity of Moroccan faba bean (Vicia faba L.) landraces should be exploited to improve the competitive ability of faba beans against weeds. The objective of this study was to in-detail evaluate competitive ability and productivity of a collection of Moroccan faba bean landraces relative to a model weed, Sinapis alba. Sixty Moroccan faba bean landraces and two checks were evaluated in 2011 and 2013 for grain yield and further agronomic traits, with and without model weed stress, under field conditions at two locations in the northwest of Morocco, using a split-plot design. The landraces showed significant genetic variation for productivity and competitive ability. Several landraces exhibited higher values compared to the two checks. Yield loss attributable to weed stress was, on average, about 69%; the weed competitive index was, on average, about 1.28. Landraces × weed treatment interaction for yield was a highly significant and marked source of variation. Several landraces were found to combine high levels of both, competitive ability against the model weed and productivity.
Key words: Vicia faba L., Sinapis alba L., Weed tolerance, Weed suppressive ability, Mean productivity
Die genetische Diversität marokkanischer Ackerbohnen (Vicia faba L.) sollte zur Verbesserung der Konkurrenzkraft von Ackerbohnen gegen Unkraut genutzt werden. Das Ziel dieser Arbeit war die detaillierte Evaluierung der Konkurrenzkraft und Produktivität einer Sammlung marokkanischer Ackerbohnen-Landrassen gegenüber dem Modellunkraut Sinapis alba. Sechzig marokkanische Ackerbohnen-Landrassen und zwei Standards wurden 2011 und 2013 auf Kornertrag und weitere agronomische Merkmale untersucht, mit und ohne Modellunkraut, in einer Spaltanlage unter Feldbedingungen an zwei Orten im Nordwesten Marokkos. Die Landrassen zeigten signifikante genetische Variation für Produktivität und Konkurrenzkraft. Mehrere Landrassen erreichten höhere Werte als die Standards. Der Ertragsverlust durch den Unkrautstress betrug im Mittel 69%, der Unkraut-Konkurrenz-Index betrug im Mittel rund 1,28. Die Landrassen × Unkraut-Interaktion für Ertrag war eine hoch signifikante und bedeutende Variationsursache. Einige Landrassen kombinierten eine hohe Konkurrenzkraft gegen das Modellunkraut mit überlegener Produktivität.
Stichwörter: Vicia faba L., Sinapis alba L., Unkrauttoleranz, Unkrautunterdrückungs-Vermögen, Mittlere Produktivität
CI – Weed competitive index; CV – Coefficient of variation; Mat – Maturity time; MP – Mean productivity; TN – Tiller number; PH – Plant height; PN(MS) – Pod number at main stem; PN(SS) – Pod number at secondary stem; PN/Pl – Pod number per plant; PS – Pod size; SN/Pl – Seed number per plant; SN/P – Seed number per pod; TSW – Thousand seed weight; Y0 – Weed–free grain yield; Y1 – Weedy grain yield; YL – Yield loss; W – Yield of white mustard; WT – Weed tolerance; WSA – Weed suppressive ability
Faba bean (Vicia faba L.) is the most important grain legume in Morocco. It covers about half of the total food legume area (191 000 ha) (MAPM, 2015). It is produced in various traditional production regions for human consumption and animal feed. Despite its importance, the cultivation of faba bean continues to decline because of low and unstable yields across environments. Both abiotic and biotic stresses are the major limiting factors. The Moroccan production of faba bean is mainly based on the exploitation of locally adapted landraces (up to 90%) in traditional farming systems. These landraces have been maintained according to farmers’ local knowledge for decades (Sadiki et al., 2002).
Faba bean production is adversely affected by weeds, especially during the early growth stages (Frenda et al., 2013). Weeds compete with the crop for limited resources, such as nutrients, water and light. They can cause heavy yield losses and decrease yield quality (Naylor, 2008). Weeds were estimated to cause seed yield reduction in faba bean of up to 60% (Frenda et al., 2013; Ghaouti et al., 2016).
Farmers in traditional agroecosystems in Morocco very rarely use chemical and/or physical control options for weed management. Weeding is very rarely practiced because of high labor costs (Alaoui, 2007). Hence, the utilization of varieties that can tolerate weeds or reduce weed growth and seed production could, therefore, be a relevant, integrated weed-management strategy (Fisher et al., 2001; Lemerle et al., 2001a). The genetic improvement of competitive ability is an attractive environment-friendly weed-management option for both low and high-input cropping systems (Cousens and Mokhtari, 1998; Dingkuhn et al., 1999).
Competitive ability of a crop in the context of weed stress has two components: (1) weed tolerance (WT), i.e., the ability to withstand the competitive impact of weeds, demonstrated by minimal yield loss, and (2) weed suppressive ability (WSA), i.e., the capacity to inhibit weed germination, growth, or reproduction (Goldberg, 1990; Jannink et al., 2000; Zhao et al., 2006a; Andrew et al., 2015). In the long term, WSA is preferred to WT, because weed suppression reduces weed seed production and their levels in the weed seed bank (Jannink et al., 2000; Zhao et al., 2006a). The ideal weed-competitive crop genotypes are able to produce high and stable yields under both weedy and weed-free conditions, and possess a strong WSA (Bussan et al., 1997).
One can select for competitive ability directly in the presence of weeds or indirectly under non-competitive conditions, based on secondary traits related to WSA. A large number of studies have been conducted to determine the competitive ability of crop species and the association of crop competitive ability with specific morphological and crop-growth traits (Place et al., 2011; Zystro et al., 2012; Korres et al., 2016). In crops such as wheat (Triticum aestivum L.) and rice (Oryza sativa L.), relationships between crop competitive ability and plant traits have been reported, e.g., plant height, tiller number, speed of development, seed size and leaflet (Lemerle et al., 2001b; Zhao et al., 2006b; Saito et al., 2010).
Variability among cultivars in their ability to compete with weeds has been reported in many crops: wheat (Lemerle et al., 1996; 2001a; 2001b), rice (Johnson et al., 1998; Caton et al., 2003; Rodenburg et al., 2009; Saito et al., 2010), corn (Zea mays L.) (Zystro et al., 2012), and soybean (Glycine max L.) (Place et al., 2011). However, very few studies have evaluated the same in faba bean (Ghaouti et al., 2016) and none has addressed the competitive ability of faba bean landraces against weeds. Most of the faba bean breeding programs have concentrated on improving yield, disease resistance, drought tolerance and resistance against broomrapes, with little consideration given to tolerance to weed stress and to crop’s ability to suppress weeds.
The Moroccan local faba bean landraces contain a great deal of genetic diversity for morphological, agronomic and physiological traits (Sadiki et al., 2002; Sadiki and El Bouhmadi, 2002; Belqadi, 2003). Therefore, the objectives of this study were (1) to evaluate the weed competitive ability of Moroccan faba bean landraces and (2) to identify superior local landraces that are suitable for direct use on farmers’ fields and/or for use in breeding programs.
The plant material used in this study included 60 faba bean (Vicia faba L.) accessions collected directly from farmers of the province of Taounate, one of the main areas of cultivation (30% of the total grain legume area) and production (35% of the total grain legume production) of faba bean in Morocco(MAPM, 2015) (Table 1). Two registered varieties (Aguadulce and Fouila Defes), commonly used by farmers in Taounate because of their relatively high level of performance (grain yield), were used as checks. The 60 faba bean accessions represented local populations; the two checks were inbred lines. White mustard (Sinapis alba L.) was used as a model weed to standardize and control the distribution of weeds in the field. This cultivated cruciferous species is related to the wild species Sinapis arvensis, which is among the major weeds in faba bean grown in Morocco (Tanji, 2001).
Table 1. Origin and seed characteristics of the 60 faba bean local landraces and the two checks (Aguadulce and Fouila Defes)
Nr | Entry | Origin | Seed color† | TSW (g) | |
Village | Commune | ||||
1 | Pop.25 | BouAouad | AïnAïcha | B | 1280 |
2 | Pop.9 | BouAouad | AïnAïcha | B | 1460 |
3 | Pop.6 | Jyahna | Bsabsa | B | 1300 |
4 | Pop.10 | Jyahna | Bsabsa | W | 1280 |
5 | Pop.14 | Jyahna | Bsabsa | B | 670 |
6 | Pop.42 | Kbib | Bsabsa | B | 1470 |
7 | Pop.57 | Kbib | Bsabsa | B | 1350 |
8 | Pop.52 | Kbib | Bsabsa | BV | 1220 |
9 | Pop.55 | Kbib | Bsabsa | B | 1570 |
10 | Pop.38 | Kbib | Bsabsa | B | 1480 |
11 | Pop.36 | Kbib | Bsabsa | W | 1240 |
12 | Pop.18 | Massakin | Bsabsa | W | 1470 |
13 | Pop.1 | Massakin | Bsabsa | W | 1220 |
14 | Pop.13 | Rkbet El Mal | Bsabsa | W | 1190 |
15 | Pop.16 | Rkbet El Mal | Bsabsa | B | 620 |
16 | Pop.12 | Houara | Mezraoua | B | 1290 |
17 | Pop.60 | Hrarcha | Mezraoua | B | 1520 |
18 | Pop.47 | Hrarcha | Mezraoua | B | 1150 |
19 | Pop.44 | Hrarcha | Mezraoua | B | 1330 |
20 | Pop.22 | Oulad El Ghoul | Mezraoua | B | 1280 |
21 | Pop.58 | Oulad El Ghoul | Mezraoua | B | 1220 |
22 | Pop.33 | Oulad El Ghoul | Mezraoua | B | 1330 |
23 | Pop.53 | Oulad El Ghoul | Mezraoua | B | 1330 |
24 | Pop.15 | Bhahda | Oulad Daoud | W | 1520 |
25 | Pop.5 | Bhahda | Oulad Daoud | B | 1570 |
26 | Pop.54 | Bhahda | Oulad Daoud | B | 1370 |
27 | Pop.56 | Bhahda | Oulad Daoud | W | 1360 |
28 | Pop.43 | Bhahda | Oulad Daoud | B | 1420 |
29 | Pop.2 | Bhahda | Oulad Daoud | B | 1560 |
30 | Pop.40 | Oulad Daoud | Oulad Daoud | B | 1400 |
31 | Pop.51 | AïnKchir | Ourtzagh | B | 1170 |
32 | Pop.31 | Bab zriba | Ourtzagh | B | 1350 |
33 | Pop.45 | Bab zriba | Ourtzagh | W | 1110 |
34 | Pop.50 | Bab zriba | Ourtzagh | W | 1260 |
35 | Pop.37 | Bab zriba | Ourtzagh | WV | 920 |
36 | Pop.21 | Bab zriba | Ourtzagh | B | 1260 |
37 | Pop.34 | Bab zriba | Ourtzagh | B | 880 |
38 | Pop.30 | Bab zriba | Ourtzagh | B | 1200 |
39 | Pop.59 | Bni Hlal | Ourtzagh | W | 1250 |
40 | Pop.26 | Bni Hlal | Ourtzagh | B | 1570 |
41 | Pop.27 | Bni Hlal | Ourtzagh | B | 1430 |
42 | Pop.4 | Bni Hlal | Ourtzagh | W | 1340 |
43 | Pop.32 | Bni Hlal | Ourtzagh | B | 1260 |
44 | Pop.39 | Bni Hlal | Ourtzagh | B | 1270 |
45 | Pop.48 | Bni Hlal | Ourtzagh | B | 1350 |
46 | Pop.3 | Bni Hlal | Ourtzagh | B | 1560 |
47 | Pop.28 | Bni Hlal | Ourtzagh | B | 1500 |
48 | Pop.49 | BniMoumen | Ourtzagh | B | 1450 |
49 | Pop.8 | BniMoumen | Ourtzagh | B | 1480 |
50 | Pop.41 | Jouidar | Ourtzagh | B | 1220 |
51 | Pop.46 | Kodya | Ourtzagh | B | 1180 |
52 | Pop.24 | Kodya | Ourtzagh | W | 1080 |
53 | Pop.29 | Kodya | Ourtzagh | B | 1070 |
54 | Pop.35 | Kodya | Ourtzagh | B | 1210 |
55 | Pop.17 | Kodya | Ourtzagh | W | 1260 |
56 | Pop.19 | Kodya | Ourtzagh | W | 1200 |
57 | Pop.7 | Mrouj | Ourtzagh | BV | 1300 |
58 | Pop.23 | Mrouj | Ourtzagh | B | 1200 |
59 | Pop.20 | Mzaourou | Ourtzagh | W | 1280 |
60 | Pop.11 | Sidi Sennoun | Ourtzagh | B | 1390 |
61 | Aguadulce | INRA Morocco | W | 1300 | |
62 | Fouila Defes | INRA Morocco | W | 490 | |
† = B-Brown seed; W-White seed; BV-Mixture of brown and violet seeds. | |||||
The experiments were conducted in four environments: in 2011 and 2013 at two locations [Loukkos (experimental station, 35°4’ N, 5°59’W) and Meknes (farmer’s field, 33°52’N, 5°32’W). The climate at the study sites was typically Mediterranean, with most of the precipitation concentrated in winter and early spring. Soil type was clay loam at Loukkos and clay at Meknes.
Experiments were conducted across the two years and at the two locations, using a split-plot design. The main plot was assigned to weed-management treatments (weedy and weed-free, i.e. with and without S. alba). Subplots were assigned to faba bean genotypes (60 local landraces and two checks). One subplot per landrace, and two subplots for each of Aguadulce and Fouila Defes were sown within the weedy as well as within the weed-free main plots. All plots were hand-weeded regularly to prevent any specific interaction with natural weeds and to establish a standardized ‘weed’ pressure in the weedy treatment. One replication (R = 1) was laid out per location and year. In addition, four plots of white mustard were sown in both main plots in each experiment (i.e., in each of the four year × location combinations).
The size of each subplot was 6.0 m² (four 3-m long rows, with an inter-row spacing of 50 cm). Mixed plots (weedy, i.e. with S. alba) and pure plots (weed-free) had a sowing density of 27 faba bean seeds/m2, with the addition of 550 seeds/m2 of white mustard in the mixed plots. First, faba bean seed was sown manually at a depth of 10 cm; white mustard was sown as second step at a depth of 2 cm in the same row. The model weed and faba bean were hence grown as additive mixture following the ideas of Snaydon (1991) and Gibson et al. (1999). The trials were sown between November and December and harvested between May and June.
The bean genotypes were evaluated for various agro-morphological traits. Measurements were taken at two levels: either for entire subplots or for five individual plants taken randomly from the two central rows of each subplot. The traits assessed on individual plants were mainly related to yield components. Table 2 gives a summary of the traits and the level of measurements.
Table 2. Traits of faba beans and of the model weed white mustard (Sinapis alba)
Traits | Details | Code | Measurement unit |
Faba bean | |||
Pod size | (1 to 9), 1 = smallest pod; 9 = biggest pod in given environment | PS | Subplot |
Plant height | Distance from soil surface to the apex (cm) | PH | Plant |
Maturity time | Number of days between sowing and maturity date (> 90% pods dry) | Mat | Subplot |
Tiller number | Average of five random plants | TN | Plant |
Pod number at main stem | Average of five random plants | PN(MS) | Plant |
Pod number at secondary stem | Average of five random plants | PN(SS) | Plant |
Pod number per plant | PN/Pl = PN(MS) + PN(SS) | PN/Pl | – |
Seed number per plant | Average of five random plants | SN/Pl | Plant |
Seed number per pod | SN/P = (SN/Pl)/(PN/Pl) | SN/P | – |
Thousand seed weight | TSW = 1000×(Yield/Pl)/(SN/Pl) (g) | TSW | – |
Grain yield | Grain yield, average of five plants per subplot (expressed as t/ha) | Yield | Subplot |
White mustard | |||
Plant height | Distance from soil surface to apex (cm) | Height | Subplot |
Grain yield | Grain yield entire subplot (as t/ha) | Wk | Subplot |
The analysis of variance (ANOVA) was performed among the 60 local populations. The two non-landrace checks were excluded from ANOVA because they may not fully represent the envisaged basic pool of local faba bean landraces. PLABSTAT v.2H (L) (Utz, 2005) was employed for statistical analysis using the following linear model:
Yijk = µ + ei + wj + (we)ij + gk + (gw)jk + (ge)ik + eijk;
where Yijk is the observation of the genotype k in environment i and the weed treatment j; μ is the general mean, ei is the environmental effect, wj is the weed treatment effect assigned to the main plot, (we)ij is the main plot error, gk is the genotype effect assigned to the subplot, (gw)kj is the interaction between the genotype k and the weed treatment j, (ge)ik is the interaction between the genotype k and the environment i, and eijk is the subplot error. The four location-year combinations were modelled as four environments. Genotypes were tested for significance against the G × E interaction. As the experiment was conducted with one replicate per environment, the single plot error variance could not be explicitly estimated. Instead, it became part of the G × W × E interaction variance. This term, given as eijk in the linear model, is the error term for testing the G × W interaction for significance.
Correlation coefficients were calculated to explore relationship between the grain yield of the 62 bean genotypes, their different indices (see below) and agromorphological traits measured in weed-free conditions, based on genotype’s means across environments. The hypothesis is that highly competitive faba beans had specific agronomic traits, which distinguished them from those that were less competitive. Taking this line of reasoning, such agronomic traits were observed under weed-free conditions.
The yield loss attributable to the competition between faba bean genotypes and the white mustard was calculated at each environment using the following equation:
YLi (%) = [(Y0k – Y1k)/Y0k] × 100 = [1 – (Y1k/Y0k)] × 100
where Y0k is the grain yield of the faba beans (k) in the weed-free condition and Y1k is the grain yield of the faba beans (k) in weedy condition.
Mean productivity (MPk) was measured in each of the four environments using the following equation:
MPk = [Y0k+ Y1k]/2
To directly compare the relative performance of the faba beans with the relative performance of the white mustard in the weedy plots, the so-called weed competitive index (CI) as used by Langeroudi and Kamkar (2009), was employed, using the following equation:
CIk = [Y1k/Ymean]/[Wk/Wmean]
where Y1k is yield of the faba bean genotype (k) under the weedy condition, Ymean is the mean yield of all the faba bean genotypes in the presence of weeds in each environment, Wk is the grain yield of white mustard, when grown mixed with the faba bean genotypes (k) and Wmean is the mean yield of white mustard in all mixed plots in each environment.
The faba bean genotypes differed significantly for all agro-morphological traits, except for tiller number (Table 3). The two weed treatments caused a significant difference for all the agro-morphological traits, and the genotype × weed interaction was significant for maturity, yield and yield components.
Table 3. Combined analysis of variance with mean squares and significance levels of the sources of variation
Source of variation | DF | PS | PH | Mat | TN | PN(MS) |
Env. (E) | 3 | 20.82** | 79865.33** | 1168.56* | 6.90 | 14.7615 |
Weed (W) | 1 | 64.52** | 13266.88** | 1484.43* | 78.20* | 185.07** |
W × E | 3 | 1.20 | 308.30* | 75.46** | 7.24** | 4.35** |
Genot. (G) | 59 | 4.82** | 161.44* | 330.83** | 0.29 | 4.88** |
G × W | 59 | 0.80 | 115.95 | 259.48** | 0.21 | 0.99** |
G × E | 177 | 1.65** | 105.87 | 10.34 | 0.23+ | 0.77** |
Error | 168§ | 0.71 | 94.22 | 11.23 | 0.18 | 0.47 |
Source of variation | PN(SS) | PN/Pl | SN/Pl | SN/P | TSW | Yield |
Env. (E) | 31.68 | 58.18 | 637.31 | 0.80 | 574.98** | 15.84 |
Weed (W) | 372.43* | 1167.69** | 18301.54** | 82.74** | 4349.37** | 493.29** |
W × E | 21.31** | 17.83** | 193.49** | 0.71** | 13.39 | 4.74** |
Genot. (G) | 3.29** | 14.16** | 83.24** | 1.38** | 250.00** | 1.38** |
G × W | 1.82** | 5.68** | 39.10** | 0.34** | 24.21** | 0.75** |
G × E | 1.33 | 2.58* | 27.95** | 0.18** | 31.88** | 0.67** |
Error | 1.10 | 1.98 | 17.13 | 0.12 | 8.13 | 0.39 |
** Significant at P< 0.01; * Significant at P< 0.05; Significant at P < 0.1; §degrees of freedom are 177–9 because of 9 missing values. | ||||||
The weedy conditions caused a decrease in all traits in the presence of the model weed Sinapis alba L. (white mustard), except for plant height, which increased (Table 4). Apart from the pod number per plant and pod number per main stem, all traits showed a wider range of variation under weedy conditions than under weed-free conditions.
Table 4. Minimum (Min), mean values (Mean), maximum (Max), least significant difference (LSD) at P = 5%, phenotypic standard deviation (SD) and coefficient of variation (CV; %) for the agro-morphological traits measured under weedy and weed-free conditions across four environments
Traits | Weed-free | Weedy | |||||||||||
Min | Mean | Max | LSD0.05 | SD | CV† | Min | Mean | Max | LSD0.05 | SD | CV† | ||
Pod size (scale 1–9) | 2.00 | 5.44 | 7.50 | 1.62 | 0.97 | 17.84 | 1.50 | 4.75 | 7.14 | 1.35 | 0.93 | 19.57 | |
Plant height (cm) | 62.4 | 76.8 | 88.1 | 12.8 | 5.2 | 6.79 | 67.5 | 87.7 | 105.6 | 14.9 | 6.9 | 7.90 | |
Maturity time (days) | 95.8 | 106.7 | 120.5 | 5.4 | 8.3 | 7.81 | 95.7 | 103.2 | 120.8 | 3.4 | 8.6 | 8.3 | |
Tiller number | – | 2.17 | – | – | – | – | – | 1.36 | – | – | – | – | |
Pod number at main stem | 2.16 | 3.19 | 9.00 | 1.25 | 1.17 | 36.70 | 1.35 | 1.96 | 4.60 | 0.79 | 0.56 | 28.59 | |
Pod number at secondary stem | 0.70 | 2.24 | 7.86 | 2.00 | 1.11 | 49.54 | 0.00 | 0.45 | 1.75 | 0.70 | 0.34 | 74.72 | |
Pod number per plant | 2.95 | 5.51 | 15.84 | 2.68 | 2.21 | 40.21 | 1.40 | 2.35 | 6.35 | 1.01 | 0.74 | 31.69 | |
Seed number per plant | 6.75 | 18.59 | 34.44 | 8.19 | 5.08 | 27.34 | 3.76 | 6.31 | 12.25 | 3.13 | 1.77 | 28.05 | |
Seed number per pod | 2.17 | 3.52 | 4.66 | 0.53 | 0.51 | 14.35 | 1.88 | 2.72 | 3.67 | 0.51 | 0.46 | 16.82 | |
1000-seed weight (g) | 567.9 | 1449.0 | 1751.4 | 194.4 | 219.8 | 15.17 | 464.5 | 1262.4 | 1475.6 | 192.0 | 196.2 | 15.5 | |
Yield (t/ha) | 1.17 | 2.88 | 4.95 | 1.24 | 0.66 | 23.00 | 0.54 | 0.88 | 1.54 | 0.45 | 0.23 | 25.78 | |
CV† = (SD/Mean) × 100 | |||||||||||||
In weedy plots, the 60 faba bean genotypes and the two checks matured, on average, 3 days earlier. They showed an increase in plant height by about 14% and a reduction in tiller number by about 37% under weedy conditions (Table 4). The pod size was reduced under weedy conditions by about 13%. Pod number per main stem, at secondary stem, and per plant decreased under weedy conditions by about 39%, 80% and 57%, respectively. Seed number per plant decreased by about 66%, seed number per pod by about 23% and TSW decreased by 13% due to presence of S. alba (Table 4).
Average grain yield decreased by 69% because of weeds. Although to different extents, the grain yield of all faba bean local landraces and the two checks decreased under weedy conditions, except for Pop.2 (Fig. 1). In weedy plots, the average grain yield varied from about 0.5 t/ha to 1.5 t/ha with an average of nearly 0.9 t/ha (Table 4). In these conditions, the lowest faba bean grain yields were recorded for Pop.10, Pop.16, Pop.13 and the check Fouila Defes. The local landraces Pop.48 and Pop.54 exhibited the highest grain yield. On the other hand, there was a greater range in grain yield in weed-free plots from about 1.2 t/ha for Pop.2 to 5.0 t/ha for Pop.38. The local landraces Pop.38, Pop.47, Pop.49, Pop.17, Pop.48 and Pop.57, had a relatively high yield both under weedy and weed-free conditions (Fig. 1).
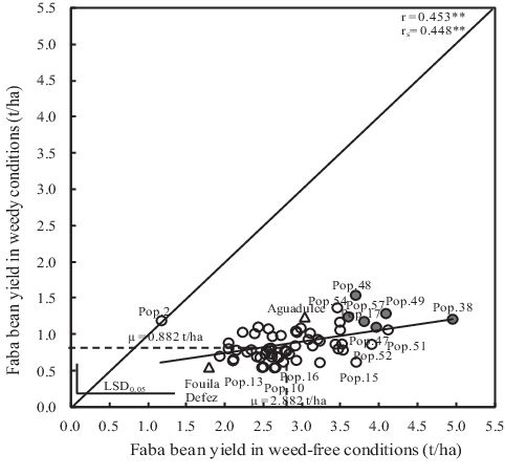
Fig. 1. Relationship of yield of the 60 faba bean local landraces and the two checks with vs. without white mustard as model weed. Symbols in grey present best genotypes in weedy and weed-free conditions (cf. text); Pearson (r) and Spearman (rS) coefficient.
Analysis of variance revealed that the faba bean genotypes differed significantly in terms of yield losses, mean productivity and weed competitive index (details not shown). The weed competitive index presented the highest range of variation (CV = 45%; Table 5).
Table 5. Minimum (Min), mean values (Mean), maximum (Max), least significant difference (LSD) at P = 5%, phenotypic standard deviation (SD) and coefficient of variation (CV; %) for yield loss, mean productivity and weed competitive index for the 60 bean local landraces and the two checks across four environments
Traits | Min | Mean | Max | LSD0.05 | SD | CV† |
Yield loss (%) | 6.61 | 69.39 | 81.38 | 17.94 | 10.84 | 16.22 |
Mean productivity (t/ha) | 1.17 | 1.89 | 3.08 | 0.71 | 0.40 | 21.08 |
Weed competitive index | 0.24 | 1.28 | 3.10 | 1.27 | 0.57 | 44.53 |
CV† = (SD/Mean) × 100 | ||||||
Using the average of the four environments, the yield losses of the 60 faba bean genotypes and the two checks varied from 7% to 81% with an average of 69% (Table 5). Pop.2 and Pop.15 obtained the lowest and the highest yield loss, respectively (Fig. 2 and 3). The average mean productivity of the 60 genotypes and the two checks in the four environments varied from about 1.2 t/ha to 3.1 t/ha with an average of about 1.9 t/ha (Table 5; Fig. 2). The check Fouila Defes presented the lowest mean productivity whereas Pop.38 had the highest mean productivity (Fig. 2).
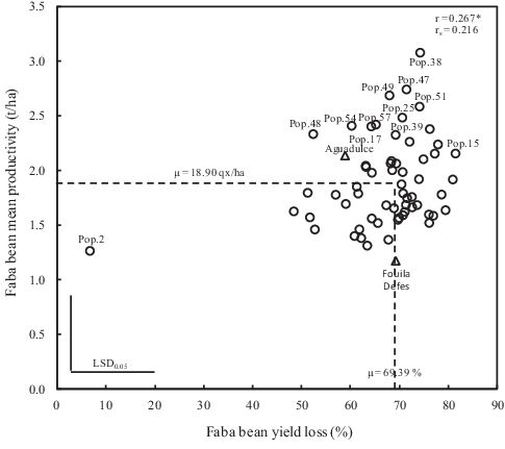
Fig. 2. Relationship between yield loss and mean productivity of the 60 faba bean local landraces and the two checks; Pearson (r) and Spearman (rS) coefficient.
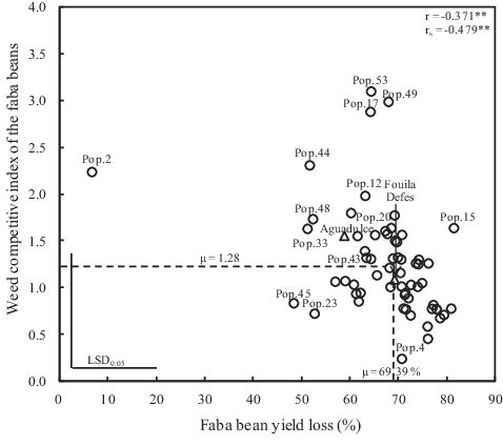
Fig. 3. Relationship between yield loss and the weed competitive index of the 60 faba bean local landraces and the two checks; Pearson (r) and Spearman (rS) coefficient.
The weed competitive index ranged from 0.24 to 3.10 with an average of 1.28 (Table 5). Pop.4 obtained the lowest weed competitive index, whereas Pop.53 presented the highest competitive index (Fig. 3), resulting from both (1) its high grain yield under weedy conditions (1.1 t/ha) compared to the average (0.9 t/ha) (Fig. 1) and (2) its pronounced ability to reduce the grain yield of white mustard from an average of 0.6 t/ha down to 0.3 t/ha; as comparison, Pop.49 achieved a similar CI, combining even higher yield under weed stress with a weaker weed suppressive ability (details not shown).
In its pure stands, the model weed S. alba grew taller (165 cm, Table 6) than all faba bean genotypes (Min: 62 cm, Max: 88 cm; Table 4). The average plant height of white mustard was lower by about 12 cm in weedy plots compared to pure stands (Table 6). Contrariwise, the average plant height of faba bean was greater by about 11 cm in weedy plots than in pure stands (Table 6). In weedy plots, white mustard’s plant height was on average 153 cm, whereas faba beans’ average plant height was 88 cm. In no case was any faba bean genotype as tall as the white mustard growing in mixture within the same plot. There were significant differences (P < 0.01) for the grain yield of S. alba grown in weedy plots depending on faba bean genotypes (details not shown). The average grain yield of white mustard in weedy plots was 0.6 t/ha and it ranged from 0.2 t/ha (with Pop.15) to 1.0 t/ha (with Pop.23; Table 6, Fig. 4). Grain yield of white mustard decreased on average by about 33% because of competition with faba bean genotypes (Table 6).
Table 6. Mean values of plant height and yield of faba bean local landraces and the two checks and white mustard in pure stand and in mixture across four environments
Trait | Height (cm) | Yield (t/ha) |
Faba bean | ||
Without white mustard (pure stand) | 76.8 a | 2.88 a |
With white mustard (mixture) | 87.7 b | 0.88 b |
White mustard | ||
Without faba bean | 165.3 a | 0.93 a |
With faba bean (mixture) | 152.9 a | 0.62 a |
Means in columns having same letter are not significantly different at 5% level of probability. | ||
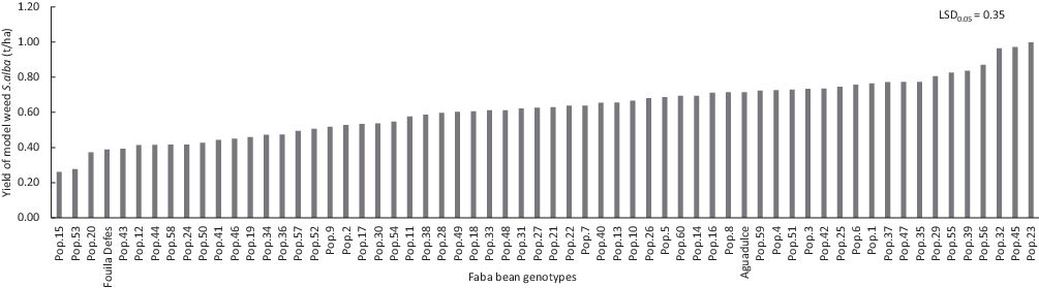
Fig. 4. Average white mustard grain yield when it was grown in mixture with one of the 60 faba bean local landraces or with one of two checks.
Grain yield in weedy stands was positively and with high significance correlated with pod size (Table 7) (r = 0.368**) and with number of seeds per pod (r = 0.397**), both measured under weed-free conditions. It was also negatively and significantly correlated with plant height (r = –0.275*), also measured under weed-free conditions. Yet, these correlations were weak. Short populations with big pods and many seeds per pod tended to yield more in the presence of the model weed S. alba.
Table 7. Coefficients of correlation among grain yield and the different indices of the 60 faba bean local landraces and the two checks and the agronomic traits measured under weed-free conditions across four environments
Traits | PS | PH | Mat | TN | PN(MS) | PN(SS) | PN/Pl | SN/Pl | SN/P | TSW | Y0 | Y1 | YL | PM | Wk |
PH | –0.177 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mat | 0.076 | 0.114 | |||||||||||||
TN | –0.132 | 0.111 | –0.123 | ||||||||||||
PN(MS) | –0.685** | 0.323* | –0.110 | 0.215 | |||||||||||
PN(SS) | –0.646** | 0.327** | –0.153 | 0.534** | 0.841** | ||||||||||
PN/Pl | –0.686** | 0.317* | –0.150 | 0.401** | 0.946** | 0.952** | |||||||||
SN/Pl | –0.343** | 0.260* | –0.061 | 0.445** | 0.738** | 0.790** | 0.838** | ||||||||
SN/P | 0.685** | –0.162 | 0.174 | –0.270* | –0.543** | –0.558** | –0.546** | –0.061 | |||||||
TSW | 0.728** | –0.239 | 0.178 | –0.161 | –0.822** | –0.752** | –0.804** | –0.585** | 0.478** | ||||||
Y0 | 0.352** | 0.075 | 0.095 | 0.391** | –0.024 | 0.137 | 0.126 | 0.570** | 0.446** | 0.299* | |||||
Y1 | 0.368** | –0.275* | –0.022 | 0.110 | –0.191 | –0.105 | –0.108 | 0.189 | 0.397** | 0.217 | 0.453** | ||||
YL | –0.008 | 0.264* | 0.111 | 0.268* | 0.153 | 0.211 | 0.213 | 0.353** | 0.044 | –0.009 | 0.435** | –0.485** | |||
PM | 0.452** | –0.003 | 0.077 | 0.355** | –0.083 | 0.076 | 0.063 | 0.518** | 0.497** | 0.300* | 0.943** | 0.631** | 0.267* | ||
Wk | 0.165 | –0.022 | 0.143 | 0.081 | 0.036 | 0.035 | 0.019 | 0.008 | 0.012 | 0.034 | –0.006 | 0.015 | 0.027 | 0.050 | |
CI | 0.140 | –0.160 | –0.183 | –0.050 | –0.091 | –0.114 | –0.057 | 0.076 | 0.197 | 0.157 | 0.226 | 0.583** | –0.371** | 0.298* | –0.550** |
Significant at ** P < 0.01 and * P < 0.05. | |||||||||||||||
A positive relationship was found between weed-free grain yield and weedy grain yield of the 62 bean genotypes (r = 0.453**). When selecting the 12 best genotypes, according to either weed-free or weedy grain yield, these two groups of selection share six common local landraces (cf. Fig. 1).
Yield loss was correlated to weed-free grain yield of the faba bean landraces and the two checks (r = 0.435**). Genotypes with good grain yield performance in weed-free conditions tended to lose more yield due to weed. Yield loss was also positively correlated with plant height (r = 0.264*), tiller number (r = 0.268*) and seeds per plant (r = 0.353**) under weed-free conditions. Tall and highly tillering faba beans tended to lose more yield in competition with S. alba.
Genotypes with high mean productivity were characterized by high yield component performances: large pods, many tillers and many seeds per plant, seeds per pod and high TSW.
The weed competitive index of faba bean landraces and the two checks was negatively correlated with faba bean yield loss (r = –0.371**) and with yield of the model weed S. alba (r = –0.550**), reflecting the algebra of this index. No relationship was found between the yield of the model weed S. alba and the other traits recorded.
This experiment was carried out in order to study the competitive ability of faba bean local landraces against white mustard (S. alba) employed as model weed. Two aspects of crop competitive ability can be defined. The first is the ability of a crop to withstand the competitive impact of weed and minimize yield loss, and the second is the ability of the crop to suppress or reduce weed growth (Goldberg, 1990; Jannink et al., 2000; Zhao et al., 2006b; Hansen et al., 2008; Andrew et al., 2015). These are described by different terms in the literature; here we describe it, respectively, as “weed tolerance” and “weed suppressive ability”.
The studied faba bean landraces differed in their weed tolerance patterns in most aspects. For plant height, all faba bean genotypes showed the same reaction to the presence of S. alba. In mixed stands, faba bean was the weaker competitor for light and its reaction to the presence of the model weed S. alba resulted in an increase in its plant height (87.7 cm > 76.8 cm; Table 6). In order to determine whether taller genotypes increased height more or less than shorter ones, a coefficient of correlation between the gain in plant height in presence of S. alba and plant height in optimal conditions was calculated and a highly significant, negative correlation was found (r = –0.540**, details not shown). Shorter genotypes tended to increase plant height under weedy conditions more than taller ones. Yet, the genotype × weed interactions failed to be significant for plant height (Table 3). As corroboration, no significant relationship was found between gain of plant height and both weedy grain yield (r = 0.121; details not shown) and yield loss (r = –0.249; details not shown). Nevertheless, tall plant height has been reported to contribute to the competitive ability in several studies (Huel and Hucl, 1996; Lemerle et al., 1996; Ogg and Seefeldt, 1999). Results in this study indicate that taller faba bean genotypes tended to yield less under weedy conditions (r = –0.275*) and to record more yield loss because of weed competition (r = 0.264*) (Table 7). This finding may be specific to the given situation that the model weed here was in all cases taller than the cultivated crop.
In the present study, maturity was not found to be an indicator of the level of tolerance to weed stress. In contrast, Huel and Hucl (1996) found that increased weed tolerance was associated with early maturity in wheat.
Tiller number of faba bean genotypes was reduced in the presence of S. alba. Such tiller number reductions have been demonstrated in wheat, barley and oat cultivars (Andrew et al., 2015). Lemerle et al. (1996) reported that high tiller number was associated with low yield reduction for wheat varieties. In the present study, no significant difference between faba bean local landraces was recorded for the number of tillers. Moreover, the genotypes did not interact significantly with the presence or the absence of weed stress for their number of tillers.
The variance of grain yield among faba bean local landraces was higher under weed-free conditions compared to weedy conditions. Indeed, the standard deviation among faba bean genotypes was 0.66 t/ha under weed-free conditions versus only 0.23 t/ha in weedy conditions; yet, the corresponding coefficients of variation were quite similar (Table 4). A smaller variation under stress is in agreement with Zhao et al. (2006b) who found such pattern for aerobic rice cultivars. Other authors observed that the genetic variation was higher under weedy than under weed-free conditions for faba bean (Ghaouti et al., 2016) and for lowland rice varieties (Rodenburg et al., 2009).
A positive relationship was observed between grain yield of faba bean local landraces and the two checks under weedy and weed-free conditions (r = 0.453**, Fig. 1). Strong positive correlations between weed-free grain yield
and weedy grain yield were observed for wheat (Cousens and Mokhtari, 1998; Lemerle et al., 2001b) and for faba bean (Ghaouti et al., 2016). Conversely, Langeroudi and Kamkar (2009) found no relationship between grain yield under weed-free and weedy conditions, when canola (Brassica napus) was grown in the presence of wild mustard (Sinapis arvensis). Although the correlation between grain yield under weedy and weed-free conditions was significant, it was not very high (r = 0.453**; Fig. 1). Highest yielding genotypes in weed-free conditions were not necessarily highest yielding in weedy conditions (e.g. Pop.52; Fig. 1).
Average yield loss ranged from 7% to 81% (Table 5). Frenda et al. (2013) reported a mean yield reduction of 60% in faba bean when it was left to compete with wild mustard and Italian ryegrass as artificial weed species. Yield loss of faba bean local landraces and the two checks was positively correlated with their yield performance in the absence of white mustard. This is in agreement with the findings of De Lucas and Froud-Williams (1994) who found that wheat varieties which produced the greatest grain yield in weed-free plots were most severely affected by weed competition. Ghaouti et al. (2016) reported that yield loss of faba bean genotypes because of the competition with weeds was negatively correlated with their yield performance in the absence of weeds. This result was clearly because of different groups of genotypes with different inbreeding levels that were collected into a joint analysis (F1 hybrids, pure lines and partly heterotic genotypes). Other authors reported that no relationship was observed between the level of grain yield in optimal conditions and yield loss for wheat (Cousens and Mokhtari, 1998; Lemerle et al., 2001b; Coleman et al., 2001) and barley (Didon and Boström, 2003).
Genotypes with high mean productivity tended to have higher yield losses in presence of the model weed S. alba (Fig. 2). Pop.2 was an outlier and the coefficient of correlation was recalculated excluding this landrace. As a result, a non-significant correlation was found (r = 0.179, details not shown). This indicates that both indices are independent and are providing complementary information on the level of mean productivity versus the level of weed tolerance. Hence, it should be possible to find genotypes that combine both high yield and high weed tolerance. Pop.48 and the variety Aguadulce correspond to this objective.
High and stable yield seems to be the priority as selection criteria (Haefele et al., 2004). However, in farmer’s fields, suppression or reduction of weed growth could be also an important characteristic (Jannink et al., 2000). Faba bean local landraces differed in their weed competitive index (CI). CI is the ratio between (1) yield of a specific bean when competing with S. alba, relative to the average of all faba beans in weedy conditions and (2) yield of S. alba when competing with this specific bean, relative to the mean of S. alba in all mixtures. CI values can be large either because a specific bean genotype yields higher than the other genotypes under weed pressure, or because it leads through its suppressive ability to a relatively low yield of S. alba in mixture, or for both effects. For instance, Pop.53 had the highest CI (3.10; Fig. 3), which is attributed to both, its higher weedy grain yield (1.05 t/ha; Fig. 1) and its ability to reduce S. alba grain yield (0.21 t/ha; Fig. 4).
No significant relationship was found between the weed-free grain yield of faba bean and the yield of white mustard, when grown in mixture (r = –0.006, Table 7). Highest yielding faba bean genotypes under weed-free conditions were not necessarily the most suppressive genotypes of S. alba. No significant relationship was found between the grain yield of the model weed S. alba in mixtures and the yield loss of faba beans (r = 0.027; Table 7). This is in agreement with the findings of Langeroudi and Kamkar (2009).
A highly significant, negative relationship was found between the yield loss and the CI (r = –0.371**; Table 7). Yet, this association is far from complete. Some faba beans (e.g. Pop.45 and Pop.23) showed in spite of low yield loss rather low CI values (Fig. 3), in these cases because these two populations allowed very high seed yield to S. alba (Fig. 4). The coefficient of correlation was recalculated between the yield loss and weed competitive index excluding the Pop.2. The coefficient of correlation remained nearly the same, yet significant only at P = 0.05 (r = –0.323*, details not shown). The negative relationship between the competitive index and yield loss seems rather to be a general trend of the studied genotypes set.
Ideal genotypes have high yield under both weed-free and weedy conditions and a distinct weed suppressive ability (Bussan et al., 1997). In the present study, Pop.57, Pop.17, Pop.34, Pop.12, Pop.36 and Pop.53, achieved a mean productivity above average, and combined this with lower than average yield loss and model weed yield, and with higher than average CI (Fig. 2, 3, and 4). These genotypes are rather competitive genotypes.
The present study did not show any significant relationship between the weed-free grain yield Y0k and the weed suppressive ability. This is true, whether the focus is on faba bean-influenced weed yield directly (Wk) or on the more complex parameter CI: r (Y0k, Wk,) = –0.006 and r (Y0k, CI) = 0.226 (Table 7). Weed-free yield was not an indicator of the suppressive ability of a genotype, corroborating Langeroudi and Kamkar (2009; canola cultivars competing with wild mustard).
A useful and marked amount of genetic diversity between faba bean local landraces was displayed. The interactions between the local landraces and the two weed treatments were highly significant. This reflects a high genetic variation for competitive ability (weed tolerance and weed suppressive ability). Several of the landraces showed higher values for mean productivity and competitive ability than the two check varieties (Aguadulce, Fouila Defes). Short plant stature tended to indicate high levels of weed tolerance, which was unexpected. In the present study, several faba bean local landraces were identified as fairly competitive. These landraces should be further studied as a promising genetic source for weed tolerance and suppressive ability. The study of competitive ability is a complex issue because the composition of natural weed populations vary over space and time and also because the weed-crop interaction is more complex than the simple interaction with our single model weed. Corresponding further studies on the competitive ability of faba bean local landraces and with pure lines bred from them should be carried out. Moreover, more environments should be employed to further increase reliability and generalizability of the findings.
We very gratefully thank all scientists and the technical team who contributed to this study. We are grateful to the farmers of Taounate for their collaboration. The study was funded by the international project MEDILEG and the national project URAC (Unités de Recherches Associées au CNRST).
Alaoui, S.B., 2007: Référentiel pour la conduite technique de la fève (Vicia faba). http://www.fellah-trade.com/ressources/pdf/feve.pdf. Access 4. 12. 2017.
Andrew, I.K.S., J. Storkey, D.L. Sparkes, 2015: A review of the potential for competitive cereal cultivars as a tool in integrated weed management. Weed Research 55, 239-248.
Belqadi, L., 2003: Diversité, conservation et valorisation des ressources génétiques marocaines de fève (Vicia faba L.). Thèse Doctorat Es-Sciences Agronomiques. Institut Agronomique et Vétérinaire Hassan II. Rabat. 137 pages.
Bussan, A.J., C.O. Burnside, J.H. Orf, E.A. Ristau, K.J. Puettmann, 1997: Field evaluation of soybean (Glycine max) genotypes for weed competitiveness. Weed Science 45, 31-37.
Caton, B.P., A.E. Cope, M. Mortimer, 2003: Growth traits of diverse rice cultivars under severe competition: implications for screening for competitiveness. Field Crops Research 83, 157-172.
Coleman, R.K., G.S. Gill, G.J. Rebetzke, 2001: Identification of quantitative trait loci for traits conferring weed competitiveness in wheat (Triticum aestivum L.). Australian Journal of Agricultural Research 52, 1235-1246.
Cousens, R.D., S. Mokhtari, 1998: Seasonal and site variability in the tolerance of wheat cultivars to interference from Lolium rigidum. Weed Research 38, 301-307.
De Lucas, C.R., R.J. Froud-Williams, 1994: The role of varietal selection for enhanced crop competitiveness in winter wheat. Aspects of Applied Biology 4, 343-350.
Didon, U.M.E., U. Boström, 2003: Growth and development of six barley (Hordeum vulgare ssp. vulgare L.) cultivars in response to a model weed (Sinapis alba L.). Journal of Agronomy and Crop Science 189, 409-417.
Dingkuhn, M., D.E, Johnson, A. Sow, A.Y. Audebert, 1999: Relationship between upland rice canopy characteristics and weed competitiveness. Field Crops Research 61, 79-95.
Fisher, A.J., H.V. Ramirez, K.D. Gibson, B.D.S. Pinheiro, 2001: Competitiveness of semidwarf upland rice cultivars against palisadegrass (Brachiaria brizantha) and signalgrass (B. decumbens). Agronomy Journal 93, 967-973.
Frenda, A.S., P. Ruisi, S. Saia, B. Frangipane, G. Di Miceli, G. Amato, D. Giambalvo, 2013: The critical period of weed control in faba bean and chickpea in Mediterranean areas. Weed Science 61, 452-459.
Ghaouti, L., A. Schierholt, W. Link, 2016: Effect of competition between Vicia faba and Camelina sativa as a model weed in breeding for organic conditions. Weed Research 56, 159-167.
Gibson, D.J., J. Connolly, D.C. Hartnett, J.D. Weidenhamer, 1999: Designs for greenhouse studies of interactions between plants. Journal of Ecology 87, 1-16.
Goldberg, D.E., 1990: Components of resource competition in plant communities. In: Grace, J., G.D. Tilman (Eds.). Perspectives in Plant Competition. Academic Press Inc., San Diego, California, USA. Pp. 27-49.
Haefele, S.M., D.E. Johnson, D.M. Bodj, M.C.S. Wopereis, K.M. Mieza, 2004: Field screening of diverse rice genotypes for weed competitiveness in irrigated lowland ecosystems. Field Crops Research 88, 39-56.
Hansen, P.K., K. Kristensen, J. Willas, 2008: A weed suppressive index for spring barley (Hordeum vulgare) varieties. Weed Research 48, 225-236.
Huel, D.G., P. Hucl, 1996: Genotypic variation for competitive ability in spring wheat. Plant Breeding, 115, 325-329.
Jannink, J.L., J.H. Orf, N.R. Jordan, R.G. Shaw, 2000: Index selection for weed suppressive ability in soybean. Crop Science 40, 1087-1094.
Johnson, D.E., M. Dinghuhn, M.P. Jones, M.C. Mahamane, 1998: The influence of rice plant type on the effect of weed competition on Oryza sativa and Oryza glaberrima. Weed Research 38, 207-216.
Korres, N.E, J.K. Norsworthy, P. Tehranchian, T.K. Gitsopoulos, D.A. Loka, D.M. Oosterhuis, D.R. Gealy, S.R. Moss, N.R. Burgos, M.R. Miller, M. Palhano, 2016: Cultivars to face climate change effects on crops and weeds: a review. Agronomy for Sustainable Development 36, 12.
Langeroudi, A.R.S., B. Kamkar, 2009: Field screening of canola (Brassica napus) cultivars against wild mustard (Sinapis arvensis) using competition indices and some empirical yield loss models in Golestan Province, Iran. Crop Protection 28, 577-582.
Lemerle, D., G.S. Gill, C.E. Murphy, S.R. Walker, C.R. Cousens, S. Mokhtari, S.J. Peltzer, R. Coleman, D.J. Luckett, 2001a: Genetic improvement and agronomy for enhanced wheat competitiveness with weeds. Journal of Agricultural Research 52, 527-548.
Lemerle, D., B. Verbeek, R.D. Cousens, N.E. Coombes, 1996: The potential for selecting wheat varieties strongly competitive against weeds. Weed Research 36, 505-513.
Lemerle, D., B. Verbeek, B. Orchard, 2001b: Ranking the ability of wheat varieties to compete with Lolium rigidum. Weed Research 41, 197-209.
MAPM, 2015: Ministère de l’agriculture et de la pêche maritime. Bilan de la campagne agricole 2013-2014. Direction de la production végétale, Département des statistiques. Rabat, Maroc. http://www.agriculture.gov.ma/. Access 4. 12. 2017.
Naylor, R.E.L., 2008: Weed Management Handbook, 9th edition. Blackwell Publishing, England.
Ogg, A.G., S.S. Seefeldt, 1999: Characterizing traits that enhance the competitiveness of winter wheat (Triticum aestivum) against jointed goatgrass (Aegilops cylindrica). Weed Science 47, 74-80.
Place, G.T., S.C. Reberg-Horton, D.A. Dickey, T.E. Carter, 2011: Identifying soybean traits of interest for weed competition. Crop Science 51, 2642-2654.
Rodenburg, J, K. Saito, R.G. Kakaï, A. Touré, M. Mariko, P. Kiepe, 2009: Weed competitiveness of the lowland rice varieties of NERICA in the southern Guinea Savanna. Field Crops Research 114, 411-418.
Sadiki, M., A. Birouk, A. Bouizzgaren, L. Belqadi, K. Rh’rrib, M. Taghouti, S. Kerfal, M. Labhlili, H. Bouhya, R. Douiden, S. Saidi, D.I. Jarvis, 2002: La diversité génétique in situ du blé dur, de l’orge, de la luzerne et de la fève: Options de stratégie pour sa conservation. In: Birouk, A., M. Sadiki, F. Nassif, S. Saidi, H. Mellas, A. Bammoun, D.I. Jarvis (Eds.) La conservation in-situ de la biodiversité agricole: un défi pour une agriculture durable. Actes du Séminaire National. 21-22 Janvier, Rabat, Maroc. IPGR, Rome. Pp. 43-121.
Sadiki, M., K. El Bouhmadi, 2002: Evaluation de populations locales marocaine de fève et de féverole pour la tolérance à la sécheresse. In: Birouk A., M. Sadiki, F. Nassif, S. Saidi, H. Mellas, A. Bammouon, D.I. Jarvis (Eds.) La conservation in-situ de la biodiversité agricole: un défi pour une agriculture durable. Actes du Séminaire National. 21-22 Janvier, Rabat, Maroc. IPGR, Rome. Pp. 242-262.
Saito, K, K. Azoma, J. Rodenburg, 2010: Plant characteristics associated with weed competitiveness of rice under upland and lowland conditions in West Africa. Field Crops Research 116, 308-317.
Snaydon, R.W., 1991: Replacement or additive designs for competition studies? Journal of Applied Ecology 28, 930-946.
Tanji, A., 2001: Adventices de la fève non irriguée dans la province de Settat. Al Awamia (Moroccan Journal of Agricultural Research) 103, 71-81.
Utz, F., 2005: PLABSTAT. Software for statistical analyses of plant breeding experiments. Version 3Awin, November 2004. University of Hohenheim, Germany.
Zhao, D.L., G.N. Atlin, L. Bastiaans, J.H.J. Spiertz, 2006a: Comparing rice germplasm groups for growth, grain yield and weed-suppressive ability under aerobic soil conditions. Weed Research 46, 444-452.
Zhao, D.L., G.N. Atlin, L. Bastiaans, J.H.J. Spiertz, 2006b: Cultivar weed-competitiveness in aerobic rice: Heritability, correlated traits, and the potential for indirect selection in weed-free environments. Crop Science 46: 372-380.
Zystro, J.P., N. De Leon, W.F. Tracy, 2012: Analysis of traits related to weed competitiveness in sweet corn (Zea mays L.). Sustainability 4, 543-560.

 Suchen
Suchen