
In situ immunofluorescence localization: A method for rapid detection of Beauveria spp. in the rhizosphere of Quercus robur saplings
In-situ-Immunfluoreszenzlokalisierung: Eine Methode zum schnellen Nachweis von Beauveria spp. in der Rhizosphäre von Quercus robur-Setzlingen
Journal für Kulturpflanzen, 71 (7). S. 211–218, 2019, ISSN 1867-0911, DOI: 10.5073/JfK.2019.07.02, Verlag Eugen Ulmer KG, Stuttgart
 This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/deed.en).
This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/deed.en).For biological control of plant pests, e.g. cockchafer grubs, in the rhizosphere of oak, apple or pine trees, entomopathogenic Beauveria spp. are increasingly applied. For successful use, it is important to monitor the spread and persistence of the inoculated fungi, both qualitatively and quantitatively. The determination of both parameters by plating on selective nutrient media or by molecular methods such as PCR of soil samples are quite laborious and often do not yield satisfactory results. Therefore, the aim of the present study was to develop a specific in situ method using immunofluorescence labelling of Beauveria spp. growing on young fine roots of three-year old oak saplings. All fine roots investigated were covered with a dense net of soil rhizosphere fungi, as visualized by staining with the nonspecific dye blankophor. On non-inoculated roots, polyclonal Beauveria antibodies did not label any of these naturally growing fungi. Only samples of roots inoculated with Beauveria brongniartii displayed specific labelling up to ten months after inoculation. Whereas the natural rhizosphere fungi were detected growing in the intercellular space of the root cortex in an ectomycorrhiza-like manner up to the endodermis, hyphae of the inoculated B. brongniartii were never seen within the root tissue but only growing on the surface of the rhizodermis. These observations indicate that B. brongniartii does not grow endophytically, and that the method used allows to discriminate B. brongniartii from the resident fungal flora in the oak tree rhizosphere. Detection by immunofluorescence labelling employed in the current study may be a useful tool to follow B. brongniartii in experiments aimed at establishing the entomopathogen in the rhizosphere and to monitor its fate in long-term control of entomopathogens.
Key words: biological pest control, blastospores, entomopathogenic fungi, immuno-fluorescence microscopy, Quercus robur, sustainability.
Zur biologischen Bekämpfung von Pflanzenschädlingen, z.B. Maikäfer-Engerlingen, in der Rhizosphäre von Eichen, Apfelbäumen oder Kiefern werden zunehmend entomopathogene Beauveria-Spezies eingesetzt. Für eine erfolgreiche Anwendung ist es wichtig, die Ausbreitung und Persistenz der ausgebrachten Pilze qualitativ und quantitativ zu erfassen. Die Bestimmung beider Größen durch Ausplattieren auf selektiven Nährmedien oder durch molekulare Methoden wie PCR ist mühsam und oft ungenügend. Ziel der vorliegenden Studie war daher, eine spezifische In-situ-Methode durch Immunfluoreszenzmarkierung von Beauveria spp. zu entwickeln, hier an jungen Feinwurzeln dreijähriger Stieleichen. Durch Anfärben mit dem unspezifischen Farbstoff Blankophor wurde sichtbar, dass alle untersuchten Feinwurzeln ein dichtes Netz von Bodenpilzen trugen. Polyklonale Beauveria-Antikörper markierten an nicht beimpften Wurzeln keinen dieser natürlich wachsenden Pilze. Mit Beauveria brongniartii beimpfte Wurzeln zeigten bis zu zehn Monate nach der Inokulation eine spezifische Markierung. Während die natürlich vorkommenden Rhizosphären-Pilze in den Interzellularräumen der Wurzelrinde wuchsen, waren Hyphen von inokulierter B. brongniartii nie im Wurzelgewebe zu finden, sondern nur oberflächlich auf der Rhizodermis. Diese Beobachtungen zeigen, dass B. brongniartii bei Eichenwurzeln nicht endophytisch wächst, und dass die verwendete Methode die Unterscheidung von B. brongniartii von der in der Eichen-Rhizosphäre lebenden Pilzflora ermöglicht. Immunfluoreszenzmarkierung, wie in der aktuellen Studie eingesetzt, kann eine nützliche Methode sein, um B. brongniartii in der Rhizosphäre nachzuweisen und quantitativ zu erfassen und somit eine Langzeitkontrolle von Schädlingen mit Entomopathogenen zu ermöglichen.
Stichwörter: Biologische Bekämpfung, Blastosporen, entomopathogene Pilze, Immunfluoreszenzmikroskopie, Quercus robur, Nachhaltigkeit.
Besides being natural enemies of a range of insects and having the ability to regulate insect populations, entomopathogenic fungi can function as antagonists of plant pathogens, beneficial rhizosphere-associates, plant growth promoters, and plant endophytes. These features are opening new possibilities in their application in integrated pest management (IPM) (Vega et al., 2009; Sánchez-Peña et al., 2011; Jaber and Ownley, 2018). Fungal endophytes are microorganisms that live for all or a portion of their life inside healthy plant tissues without causing any apparent symptoms of disease (De Bary, 1884; Wilson, 1995; Vega et al., 2008; Greenfield et al., 2016).
The entomopathogenic fungus Beauveria bassiana (Balsamo) Vuillemin (Hypocreales: Clavicipitaceae) occurs in terrestrial ecosystems worldwide (Bidochka et al., 1998; Meyling and Eilenberg, 2007; McKinnon et al., 2018) and has the ability to colonize plants naturally, or after artificial inoculation (Wagner and Lewis, 2000; Vidal and Jaber, 2015; Rondot and Reineke, 2018). Using molecular methods, B. bassiana has been recorded as an endophyte in a range of plant species (Posada et al., 2007; Reay et al., 2010; Biswas et al., 2015). Beauveria brongniartii (Sacc.) Petch is a soil borne entomopathogenic fungus that is widespread in soils. Certain strains of B. brongniartii cause significant mortality in various insect pests and have potential as biological control agents (Jones, 1994; Keller et al., 1999; Kheswa, 2016). Pedunculate oak (Quercus robur L.) is one of the most valuable tree species in Croatia and very susceptible towards adverse abiotic and biotic environmental factors, such as air and water pollution, site conditions, climatic extremes, attack by different insect pests, infection by pathogenic fungi, as well as its permanent exploitation due to its commercial value (Tikvić et al., 2009). The aim of the present research was to investigate the possible endophytic and rhizospheric growth of B. bassiana and B. brongniartii in pedunculate oak leaves and roots. The focus was on the rapid and specific in situ detection of these fungi in rhizosphere soil samples by means of specific polyclonal antibodies.
Entomopathogenic fungal strains were obtained from the culture collection of the Institute for Biological Control, Julius Kühn-Institut, Darmstadt, Germany. These were Beauveria bassiana strain ATTC 74040 (re-isolate of the product NATURALIS®) and Beauveria brongniartii, JKI-BI 1249. The fungi were cultivated on potato dextrose agar (PDA, Carl Roth GmbH, Germany) at 25 ± 2°C. Blastospore suspensions were prepared by inoculation of potato glucose broth (PDB, Carl Roth GmbH, Germany) with conidia of B. bassiana or B. brongniartii. The suspensions were incubated in a rotary shaker for four days at 25°C and 150 rpm, then filtered through cotton gauze (Mullro®), and the blastospore concentration was adjusted to a concentration of 107 per ml. After confirming the spore viability with 0.01% acridine orange under blue fluorescent light (Fig. 1a, b), the suspensions were used to inoculate potted three-year-old Quercus robur L. saplings.
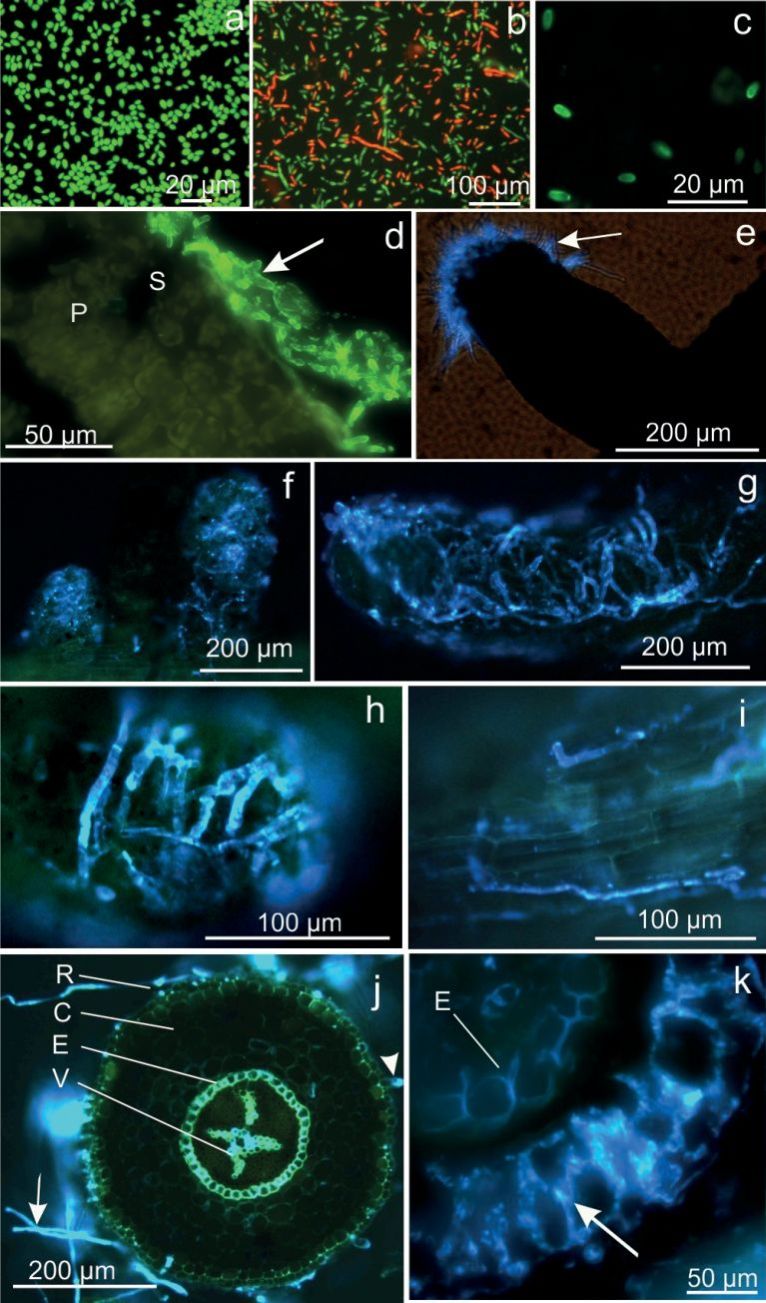
Fig. 1. Beauveria spp. blastospores and oak fine roots naturally colonized by rhizosphere fungi. a, b) Viability test of B. bassiana blastospores in culture with 0.01% acridine orange. a) Vital green fluorescent spores; b) predominantly dead red fluorescent spores. c) Beauveria bassiana spores subjected to in situ immunofluorescent specific Beauveria polyclonal antibodies/goat anti-rabbit secondary FITC-conjugated antibodies. d) Cross section of oak leaf inoculated with B. bassiana, 10 dpi, and subjected to full immunolabelling: spores and hyphae (arrow) remain outside on the epidermal leaf surface and are absent in the mesophyll, (P) palisade cells, (S) spongy mesophyll (CLSM). e-k) Fine roots of non-inoculated three-year-old control oak saplings, naturally colonized internally and/or on the surface by natural rhizospheric fungi; blankophor (0.01%) staining. e) Soil fungi colonizing preferentially the tip of fine roots (arrow). f) Series of fine roots covered with a fungal net. g) Dense fungal net around fine root tip. h) Close-up view of fungal hyphae. i) Fungi growing longitudinally along fine root tip. j) Cross section of fine root with fungal hyphae (arrow) surrounding the rhizodermis (R), (C) cortex, root hair (arrowhead), (E) endodermis, (V) vascular bundle. k) Cross section detail revealing mycorrhiza-like fungal growth in the intercellular space (arrow) up to the endodermis with Casparian strips (E). The central cylinder is free of hyphae (CLSM).
Q. robur saplings, obtained from “die-forstpflanze.de” and growing in standard steamed potting substrate (Frustorfer Erde Typ LD 80, HAWITA Gruppe GmbH, Vechta, Germany), were inoculated by dipping the leaves into B. bassiana suspensions or by tissue infiltration by gently pressing the open end of a 5 ml syringe on the lower leaf side through the stomata. For gentle infiltration, smaller plants were turned upside down with leaves submerged in the spore suspension within a beaker inside a desiccator, connected to a vacuum pump. Control plants were infiltrated with tap water. Roots of separate plants were inoculated by drenching the pots with 100 ml of B. brongniartii spore suspension. Control plants were treated with 100 ml of PDB. All plants were maintained under humid conditions for three days, and then kept in the greenhouse at 21–22°C, 60–80% RH with additional light from sodium vapor lamps (PLANTASTAR®, 400 W, NDL, 725 μmol s–1 photons, OSRAM) at a 16:8 h photoperiod. The first examination of inoculated leaves and roots started after five days, and was repeated up to ten months.
Rhizosphere fungi were non-specifically stained by submerging the samples in 0.01% blankophor (4,4'-bis-([4-(p-sulfonanilino)-6-diethanolamin-1,3,5-triazin-2-yl]amino)-stilben-2,2'-disulfonic acid; Bayer, Leverkusen, Germany), which stains 1,4 linked polymers (cellulose, chitin), for 1.5 min. For specific detection of Beauveria spp., tissue samples were subjected to immunofluorescence labelling with rabbit primary polyclonal and FITC conjugated secondary fluorescent goat anti-rabbit antibodies of B. bassiana, prepared as previously reported in detail (Ullrich et al., 2017). Randomly chosen leaf and root samples from treated plants were cut into 5 mm pieces and kept overnight at 5°C in 4% paraformaldehyde (PFA) for fixation. After thoroughly washing in PBS, the tissue pieces were dehydrated and embedded into Steedman's wax according to the method previously described in detail (Ullrich et al., 2017; according to Vitha et al., 1997). Wax samples were cut into 10–12 μm thin sections with a microtome (Cryocut CM 3050, Leica, Germany) and collected on albumin-glycerol coated slides. To block unspecific binding sites, the de-waxed sections were incubated in 1% BSA (Aurion) in 0.1x phosphate-buffered saline (PBS) for 1.5 h at 22°C. For immunolabelling, sections were incubated overnight with a rabbit anti-B. bassiana primary antibody IgG, diluted 1:200. After washing the sections, fluorescent FITC conjugate goat anti-rabbit secondary antibody (Molecular Probes, Göttingen, Germany), diluted 1:200, was applied for 2 h at 22°C. After washing, the sections were mounted on slides in glycerol containing 2.5% of the anti-fading agent 1.4-diazabicyclo[2.2.2]octane (DABCO). Generally, sections were always labelled and stained after cutting the leaves or roots.
Due to cross-reactivity, the anti-B. bassiana antibody detected further entomopathogens which all belong to the order Hypocreales (Ullrich et al., 2017). The cross-reactivity of the purified rabbit IgGs was confirmed by plate trapped antigen ELISA (PTA-ELISA) (Rohde and Rabenstein, 2005) and western blotting experiments. No reaction was observed with bovine serum albumin as negative control in PTA-ELISA. In western blotting experiments, the main reactive bands of B. bassiana and Isaria fumosorosea antigen preparations revealed an apparent molecular weight of approximately 27 or 30 kDa, respectively (data not shown). The observed immunological cross-reactivity of antibodies seems to be universal experience in fungus serology (Kaufman and Standard, 1987; Notermans et al., 1998) including plant invading fungi (Dewey, 2002). It suggests that the fungi in question share common antigenic determinants, which may demonstrate phylogenetic relationships especially with related fungi but also, to a lesser extent, with distantly related fungi (Schmechel et al., 2006; Thornton and Wills, 2015).
Using inoculated samples of oak roots (see “Inoculation of oak saplings” above) an alternative rapid method of immunofluorescence labelling for rapid visualization of B. brongniartii was developed. Roots from inoculated and non-inoculated control oak saplings were gently washed several times with tap water to remove the soil, cut into 5 mm pieces and kept in 4% PFA in glass vials at 5°C. After washing, the root pieces were incubated without further cutting in anti-B. bassiana primary antibody, diluted 1:200 in PBS for 1 h at room temperature and then at 5°C overnight. The following day they were washed, and incubated in goat anti-rabbit secondary antibody solution, diluted 1:200, and left for 2 hours at room temperature. Finally, the root samples were washed in PBS and kept in the anti-fading DABCO solution. Not only fungal hyphae were labelled by this rapid method but also the blastospores (Fig. 1 c, d).
The samples were observed under an Aristoplan epifluorescence microscope (Leica, Wetzlar, Germany) and photographed with a CCD camera (ColorView II, Olympus) using the software AnalySIS FIVE, or examined under the confocal laser scanning microscope (Leica TCS SP8 confocal microscope, Germany). CLSM images were taken by using the Leica Application Suite X (LAS X) imaging software platform for confocal microscopy. Samples stained with acridine orange and FITC were examined under fluorescent blue light (excitation at 450–490 nm), and those stained with blankophor under UV-light (excitation at 340–380 nm). For comparison, some samples were examined under differential interference contrast optics (DIC). More than 350 preparations with more than 5000 sections or tissue pieces were investigated.
Microscopical analysis of leaf and root preparations of in each case separately inoculated plants revealed that B. bassiana and B. brongniartii did not colonize oak leaf or root interior parenchyma. In the leaf sections, spores and growing hyphae were only seen on the leaf surface (Fig. 1d).
In the root samples of both inoculated and control plants, the fine roots were covered with a dense fungal net (Fig. 1f, g, h), preferentially at the tip of the fine roots (Fig. 1e), but also at young secondary roots in longitudinal wrinkles (Fig. 1i). Natural rhizospheric fungal growth in an ectomycorrhiza-like manner was present in the intercellular space of the root cortex up to the endodermis (Fig. 1j, k). The latter apparently functions as an efficient barrier against fungal invasion of the root vascular system.
Up to ten months after inoculation, germinated and established B. brongniartii hyphae were found on the root surface, especially on young fine roots (Fig. 2a-e), by following the rapid method of in situ immunofluorescence labelling of Beauveria spp. (Fig. 2c). However, in contrast to the natural rhizospheric fungi, Beauveria never invaded the oak root cortex across the cork cells of the periderm, as found in sections immunolabelled after cutting (Fig. 2f, g).
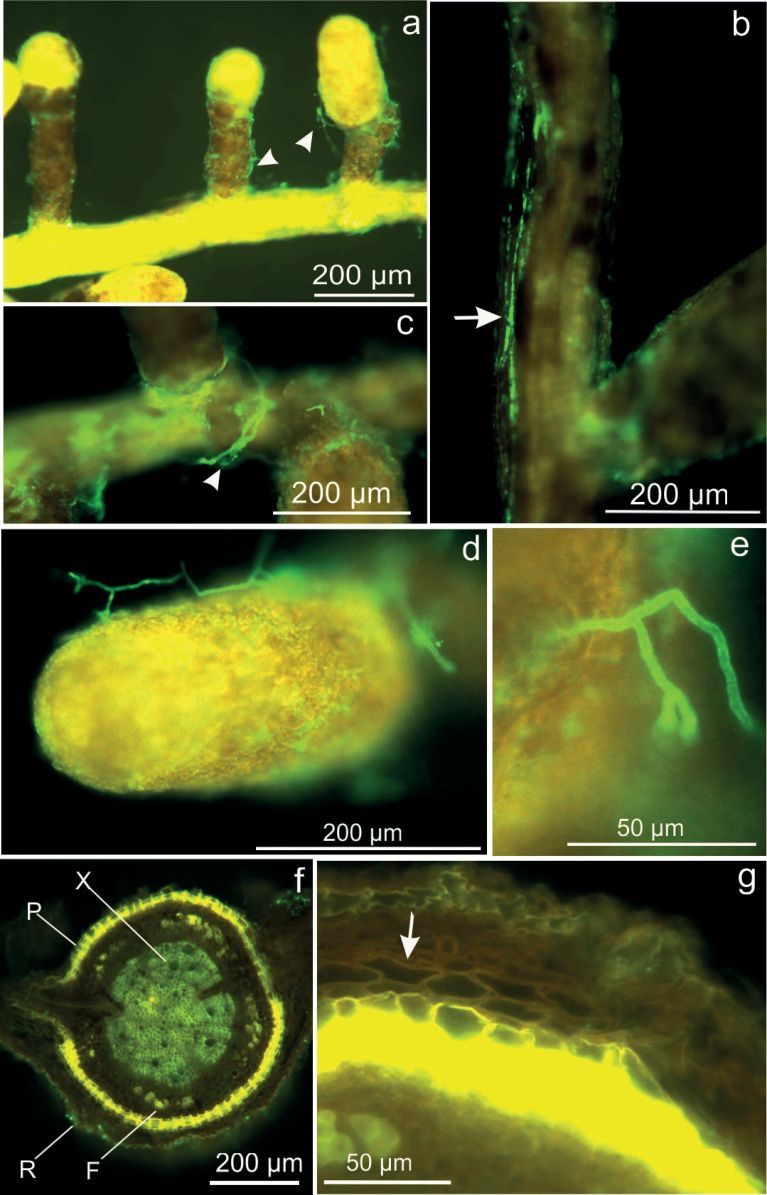
Fig. 2. Selective rapid immunofluorescent labelling of Beauveria brongniartii on oak fine roots. a) B. brongniartii green fluorescent hyphae, 5 mpi, at a series of yellow/brown fine roots (arrowheads). b) Longitudinally growing hyphae (arrow), 5 mpi. c) Hyphae surrounding the base of fine root (arrowhead), 5 mpi. d, e) Close-up view of hyphae at fine root, 5 mpi. f) Oak fine root cross section with suberinized cork cells of the periderm (P), (R) rhizodermis, (F) fibers, (X) xylem with lignified pith, and g) detail revealing absence of endophytic B. brongniartii growth inside the complete root tissue (arrow), 6.5 mpi.
After subjecting fine root samples from non-inoculated oak tree rhizosphere to immunolabelling, no hyphae were visible under blue light FITC excitation, whereas many hyphae were visible under DIC optics. This observation confirms the specificity of the immunolabelling procedure and strongly suggests that only natural rhizospheric fungi were present in the non-inoculated oak rhizosphere (Fig. 3a-d).
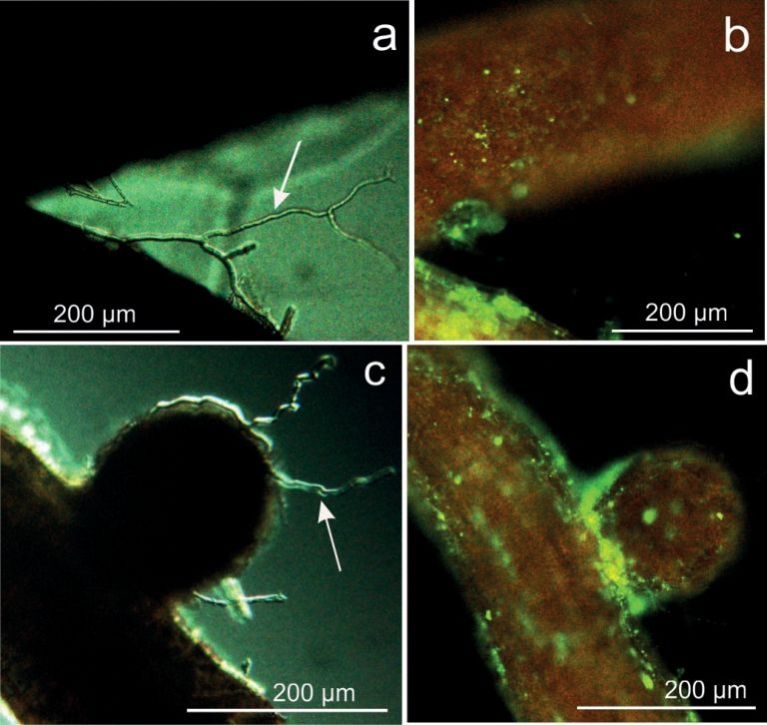
Fig. 3. Non-inoculated control oak fine roots after immunologically staining with antibodies. a, c) Differential interference contrast (DIC) micrographs showing fungal hyphae. b, d) The same samples under blue light excitation. Fluorescing hyphae are absent, which proves the selectivity of the method: immunolabelling with specific antibodies only reacts with Beauveria species, naturally occurring fungi are not detected.
Three findings of this study can be highlighted. (1.) Entomopathogenic B. bassiana and B. brongniartii did not establish as endophytes in oak tree tissues, as they were not able to grow within the leaves or roots, but only on the leaf and root surface. On the contrary, hyphae of natural rhizosphere fungi were observed growing intercellularly in the root cortex. However, growth of these fungi never extended beyond the endodermis, which is known to function as an efficient barrier against invasion of the stele also by ecto- and endomycorrhizal fungi and other fungal root endophytes (Sieber and Grünig, 2013). (2.) The method of immunofluorescence labelling of pieces of leaf and root, after fixation with PFA, turned out to be an easier and faster way to detect entomopathogenic B. bassiana and B. brongniartii on the fine root surface than the use of classical microtome sections. (3.) The fully developed hyphae of B. brongniartii that were observed surrounding the oak fine roots may indicate a commensal symbiotic relationship between plant and fungus. Their presence even ten months after inoculation apparently demonstrated a sustainable persistence of B. brongniartii in the rhizosphere.
(1.) Microscopical observation of oak leaf and root sections showed no hyphal growth of B. bassiana and B. brongniartii inside of the plant organs. Several inoculation methods were used, but none of them resulted in successful endophytic establishment of the strains of B. bassiana and B. brongniartii tested. In almost all of around 5000 sections examined by staining with dyes or labelling with antibodies, fungal spores were seen to germinate only on leaf and root surfaces, and no hyphae were observed inside the tissues. As reported previously, this could be due to several reasons (Ullrich et al., 2017; Koch et al., 2018): a. lack of enzymes of these entomopathogenic fungi able to degrade plant cell walls and cell membranes; b. lack in the ability of the fungi to acquire nutrients from the plant intercellular space; c. production of plant secondary metabolites that inhibit the spore germination, which appears to be an antifungal defense response of plants (Peng and Kuc, 1992; Bednarek et al., 2009). In addition, our results indicate that in oak rootlets the cork layers of the periderm function as an efficient barrier (Fig. 2f, g), that cannot be overcome by Beauveria spp. hyphae. By contrast, natural rhizospheric fungi appear to be able to cross this barrier (Fig. 1j, k).
(2.) In previous studies, traditional staining techniques proved valuable for the detection of entomopathogenic fungi in plants (Ullrich et al., 2017; Koch et al., 2018). Nevertheless, in the present work, immunofluorescence labelling with rabbit primary polyclonal IgG and FITC conjugated secondary fluorescent goat anti-rabbit antibodies was found to specifically detect the selected Beauveria strains. Whereas immunofluorescence labelling using traditional micro-sections is rather laborious, the method used here avoids the time consuming steps of dehydration of the tissues in graded series of ethanol, embedding in Steedman’s wax and sectioning with a microtome. Since this shorter method reliably detect the Beauveria hyphae on the surface of oak roots, it is regarded as a promising tool for further investigations. Although still rarely used at present (McKinnon et al., 2017; Card, 2018), fluorescence and confocal laser scanning microscopy are valuable tools to elucidate the interaction between endophytes and their hosts in general. The method of immunofluorescence labelling applied in the present study has the potential to complement and significantly facilitate and improve such studies, as PCR techniques do not show where and in which amount endophytes are established.
Finally, germinated B. brongniartii hyphae were found, surrounding the oak fine root surface up to ten months after inoculation, proving the ability of the fungus to persist in soil for a longer period. This is in agreement with previous observations regarding the persistence of Beauveria spp. in soil. In one study, Beauveria strains were still detected five months after application (Coombes et al., 2016). Other studies report fungal persistence in field soils of the valley of Aosta (Italy) for more than 24 months (Dolci et al., 2006), and reisolation of B. brongniartii even 14 and 15 years, respectively, after the last treatment in Switzerland and in Tyrol (Austria) (Enkerli et al., 2004; Mayerhofer et al., 2015). Those studies demonstrated the efficacy of Beauveria against different insects with soil-dwelling life stages where application of Beauveria caused a reduction of their population density. Persistence of Beauveria spp. in close association with plant roots over long periods may also indicate a balanced relationship between host and fungus. This might be a good basis for further research on the regulation of insect populations by rhizospheric growth of our B. brongniartii strain.
(3.) The results obtained in this study could indicate a kind of commensal symbiosis between oak roots and B. brongniartii hyphae that benefits the plant without negative effects on the fungus. Rather, the fungi seem to get nutrients from root exudates as they are growing closely to the germinating fine roots. This was also shown by Zitlalpopoca-Hernandez et al. (2017) with the roots of maize. A competition between native arbuscular mycorrhizal fungi (AMF) and B. bassiana was detected. However, more and quantitative studies are required to confirm that the present strain has sustainable rhizosphere colonization potential and is a possible agent for microbial pest control.
This study was supported by a scholarship of the “Deutscher Akademischer Austauschdienst” (DAAD) to MM. We thank Natascha Heil for valuable technical assistance.
Bednarek, P., M. Pislewska-Bednarek, A. Svatos, B. Schneider, J. Doubsky, M. Mansurova, M. Humphry, C. Consonni, R. Panstruga, A. Sanchez-Vallet, A. Molina, P. Schulze-Lefert, 2009: A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323, 101-106, DOI: 10.1126/science.1163732.
Bidochka, M.J., J.E. Kasperski, G.A.M. Wild, 1998: Occurrence of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana in soils from temperate and near- northern habitats. Canadian Journal of Botany 76, 1198–1204, DOI: 10.1139/b98-115.
Biswas, C., P. Dey, B.S. Gotyal, S. Satpathy, 2015: A method of multiplex PCR for detection of field released Beauveria bassiana, a fungal entomopathogen applied for pest management in jute (Corchorus olitorius). World Journal of Microbiology and Biotechnology 31, 675–679, DOI: 10.1007/s11274-015-1821-6.
Card, S., 2018: Recension to: Koch, E., P. Zink, C.I. Ullrich, R.G. Kleespies, 2018: Light microscopic studies on the development of Beauveria bassiana and other putative endophytes in leaf tissues. Journal of cultivated plants. 70, 95-107, Journal of Cultivated Plants 70, 110-111, DOI: 10.1399/JFK.2018.03.02.r1.
Coombes, C.A., M.P. Hill, S.D. Moore, J.F. Dames, 2016: Entomopathogenic fungi as control agents of Thaumatotibia leucotreta in citrus orchards: field efficacy and persistence. BioControl 61, 729–739, DOI: 10.1007/s10526-016-9756-x.
De Bary, A., 1884: Vergleichende Morphologie und Biologie der Pilze, Mycetozoen und Bacterien. Leipzig, Berlin, Verlag von Wilhelm Engelmann, DOI: 10.5962/bhl.title.42380.
Dewey, F.M., 2002: Immunological techniques. In: Waller, J.M., J.M. Lennee, S.J. Waller (eds.): Plant Pathologists Pocket Book. 3rd ed., Chapter 22, CABI Biosciences, Wallingford, Oxford, UK, pp. 221-228, ISBN 0-85199-459-8.
Dolci, P., F. Guglielmo, F. Secchi, O.I. Ozino, 2006: Persistence and efficacy of Beauveria brongniartii strains applied as biocontrol agents against Melolontha melolontha in the Valley of Aosta (northwest Italy). Journal of Applied Microbiology 100, 1063-1072, DOI: 10.1111/j.1365-2672.2006.02808.x.
Enkerli, J., F. Widmer, S. Keller, 2004: Long-term field persistence of Beauveria brongniartii strains applied as biocontrol agents against European cockchafer larvae in Switzerland. Biological Control 29, 115–123, DOI: 10.1016/S1049-9644(03)00131-2.
Greenfield, M., M.I. Gómez-Jiménez, V. Ortiz, F.E. Vega, M. Kramer, S. Parsa, 2016: Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biological Control 95, 40-48, DOI: 10.1016/j.biocontrol.2016.01.002.
Jaber, L.R., B.H. Ownley, 2018: Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biological Control 116, 36-45, DOI: 10.1016/j.biocontrol.2017.01.018.
Jones, K.D., 1994: Aspects of the biology and biological control of the European corn borer in North Carolina. Ph.D. thesis, North Carolina State University.
Kaufman, L., P.G. Standard, 1987: Specific and rapid identification of medically important fungi by exoantigen detection. Ann. Rev. Microbiol. 41, 209-225, DOI: 10.1146/annurev.mi.41.100187.001233.
Keller, S., C. Schweizer, P. Shah, 1999: Differential susceptibility of two Melolontha populations to infections by the fungus Beauveria brongniartii. Biocontrol Science and Technology 9, 441-446, DOI: 10.1080/09583159929703.
Kheswa, N., 2016: Development of Beauveria brongniartii as a bio-insecticide to control white grub (Coleoptera: Scarabaeidae) species attacking sugarcane in South Africa. Master Thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa; 167 pp, URI: http://hdl.handle.net/10413/14663.
Koch, E., P. Zink, C.I. Ullrich, R.G. Kleespies, 2018: Light microscopic studies on the development of Beauveria bassiana and other putative endophytes in leaf tissues. Journal of Cultivated Plants 70, 94-106 DOI: 10.1399/JKI.2018.03.02.
Mayerhofer, J., J. Enkerli, R. Zelger, H. Strasser, 2015: Biological control of the European cockchafer: persistence of Beauveria brongniartii after long-term applications in the Euroregion Tyrol. BioControl 60 (5), 617-629, DOI: 10.1007/s10526-015-9671-6.
McKinnon, A.C., S. Saari, M.E. Moran-Diez, N.V. Meyling, M. Raad, T.R. Glare, 2017: Beauveria bassiana as an endophyte: a critical review on associated methodology and biocontrol potential. BioControl 62, 1-17, DOI: 10.1007/s10526-016-9769-5.
McKinnon, A.C.,T.R. Glare, H.J. Ridgway, A. Mendoza-Mendoza, A. Holyoake, W.K. Godsoe, J.L. Bufford, 2018: Detection of the entomopathogenic fungus Beauveria bassiana in the rhizosphere of wound-stressed Zea mays plants. Frontiers in Microbiology 9, DOI: 10.3389/fmicb.2018.01161.
Meyling, N.V., J. Eilenberg, 2007: Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: potential for conservation biological control. Biological Control 43, 145-155, DOI: 10.1016/j.biocontrol.2007.07.007.
Notermans, S.H.W., M.A. Cousin, G.A. De Ruiter, F.M. Rombouts, 1998: Fungal immunotaxonomy. In: Frisvad, J.C., P.D. Bridge, D.K. Arora (eds.): Chemical Fungal Taxonomy. New York: Marcel Dekker, pp. 121-152, ISBN 0 8247 0069 4.
Peng, M., J. Kuc, 1992: Peroxidase-generated hydrogen peroxide as a source of antifungal activity in vitro and on tobacco leaf discs. Phytopathology 82, 696-699, DOI: 10.1094/Phyto-82-696.
Posada, F., M.C. Aime, S.W. Peterson, S.A. Rehner, F.E. Vega, 2007: Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycological Research 111, 748-757, DOI: 10.1016/j.mycres.2007.03.006.
Reay, S.D., M. Brownbridge, B. Gicquel, N.J. Cummings, T.L. Nelson, 2010: Isolation and characterization of endophytic Beauveria spp. (Ascomycota: Hypocreales) from Pinus radiata in New Zealand forests. Biological Control 54, 52–60, DOI: 10.1016/j.biocontrol.2010.03.002.
Rohde, S., F. Rabenstein, 2005: Standardization of an indirect PTA-ELISA for detection of Fusarium spp. in infected grains. Mycotoxin Res. 21, 100-104, DOI: 10.1007/BF02954429.
Rondot, Y., A. Reineke, 2018: Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insects. Biological Control 116, 82-89, DOI: 10.1016/j.biocontrol.2016.10.006.
Sánchez-Peña, S.R., J.S.-J. Lara, R.F. Medina, 2011: Occurrence of entomopathogenic fungi from agricultural and natural ecosystems in Saltillo, México, and their virulence towards thrips and whiteflies. Journal of Insect Science 11, 1-10, DOI: 10.1673/031.011.0101.
Schmechel, D., J.P. Simpson, D. Beezhold, D.M. Lewis, 2006: The development of species-specific immunodiagnostics for Stachybotrys chartarum: the role of cross-reactivity. J. Immunol. Methods 309, 150-159, DOI: 10.1016/j.jim.2005.12.001.
Sieber, T.N., C.R. Grünig, 2013: Fungal root endophytes. In: Eshel, A., T. Beeckman (eds.): The hidden half, Chapter 38, Publisher CRC Press, Taylor and Travis Group, Boca Raton, USA., pp. 31-38, DOI: 10.1201/b14550-45.
Tikvić, I., Ž. Zečić, D. Ugarković, D. Posarić, 2009: Oštećenost stabala i kakvoća drvnih sortimenata hrasta lužnjaka na spačvanskom području. Šumarski list 133, 237-248, URI: https://hrcak.srce.hr/39461.
Thornton, C.R., O.E. Wills, 2015: Immunodetection of fungal and oomycete pathogens: Established and emerging threats to human health, animal welfare and global food security. Crit. Rev. Microbiol. 41, 27-51, DOI: 10.3109/1040841X.2013.788995.
Ullrich, C.I., E. Koch, C. Matecki, J. Schäfer, T. Burkl, F. Rabenstein, R.G. Kleespies, 2017: Detection and growth of endophytic entomopathogenic fungi in dicot crop plants. Journal of Cultivated Plants 69, 291-302, DOI: 10.1399/JfK.2017.09.02.
Vega, F.E., F. Posada, M.C. Aime, M. Pava-Ripoll, F. Infante, S.A. Rehner, 2008: Entomopathogenic fungal endophytes. Biological Control 46, 72-82, DOI: 10.1016/j.biocontrol.2008.01.008.
Vega, F.E., M.S. Goettel, M. Blackwell, D. Chandler, M.A. Jackson, S. Keller, M. Koike, N.K. Maniania, A. Monzón, B.H. Ownley, J.K. Pell, D.E.N. Rangel, H.E. Roy, 2009: Fungal entomopathogens: new insights on their ecology. Fungal Ecology 2, 149-159, DOI: 10.1016/j.funeco.2009.05.001.
Vidal, S., L.R. Jaber, 2015: Entomopathogenic fungi as endophytes: plant-endophyte-herbivore interactions and prospects for use in biological control. Current Science 109, 46-54.
Vitha, S., F. Baluška, M. Mews, D. Volkmann, 1997: Immunofluorescence detection of F-actin on low melting point wax sections from plant tissues. Journal of Histochemistry and Cytochemistry 45, 89-95, DOI: 10.1177/002215549704500112.
Wagner, B.L., L.C. Lewis, 2000: Colonization of corn, Zea mays, by the entomopathogenic fungus Beauveria bassiana. Applied and Environmental Microbiology 66, 3468-3473, DOI: 10.1128/AEM.66.8.3468-3473.2000.
Wilson, D., 1995: Endophyte: the evolution of a term, and clarification of its use and definition. Oikos 73, 274–276, DOI: 10.2307/3545919.
Zitlalpopoca-Hernandez, G., M.B. Najera-Rincon, E. del-Val, A. Alarcon, T. Jackson, J. Larsen, 2017: Multitrophic interactions between maize mycorrhizas, the root feeding insect Phyllophaga vetula and the entomopathogenic fungus Beauveria bassiana. Applied soil ecology, 115, 38-43, DOI: 10.1016/j.apsoil.2017.03.014.

 Suchen
Suchen