
Differenzialdiagnose der Pathogene Pseudomonas syringae pv. aesculi und Phytophthora spp. an Aesculus spp. im Hamburger Stadtgebiet
Differential diagnosis of the pathogens Pseudomonas syringae pv. aesculi and Phytophthora spp. at Aesculus spp. in the urban area of Hamburg
Journal für Kulturpflanzen, 71 (10). S. 253–263, 2019, ISSN 1867-0911, DOI: 10.5073/JfK.2019.10.01, Verlag Eugen Ulmer KG, Stuttgart
 Dies ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Namensnennung 4.0 International Lizenz (CC BY 4.0) zur Verfügung gestellt wird (https://creativecommons.org/licenses/by/4.0/deed.de).
Dies ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Namensnennung 4.0 International Lizenz (CC BY 4.0) zur Verfügung gestellt wird (https://creativecommons.org/licenses/by/4.0/deed.de).Während der dritten Reihenuntersuchung in der Freien und Hansestadt Hamburg an Aesculus spp. wurden 94 Rosskastanien der Arten A. hippocastanum und A. carnea auf die Phytopathogene Pseudomonas syringae pv. aesculi und Phytophthora spp. untersucht. Die Bäume wurden gezielt nach dem makroskopisch erfassten Schadbild und der räumlichen Verteilung im Stadtgebiet ausgewählt. Das Verfahren der Probenahme wurde minimalst invasiv durchgeführt. Die molekularbiologische Untersuchung erfolgte am Tag der Probenahme, sodass bereits am Folgetag die Laborbefunde vorlagen. Insgesamt wurden 82 Rosskastanien positiv auf P. syringae pv. aesculi und 7 Rosskastanien positiv auf Phytophthora spp. getestet. Die Sequenzierung ergab, dass es sich bei dem Phytophthora-Erreger um Phytophthora cactorum handelt. Zusätzlich wurde an 3 Rosskastanien eine Koinfektion festgestellt.
Stichwörter: Pseudomonas syringae pv. aesculi, Phytophthora cactorum, Aesculus hippocastanum, Aesculus carnea, PCR, Hamburg, Rindenkrankheit
During the third survey in the Free and Hanseatic City of Hamburg on Aesculus spp. 94 horse chestnut trees of the species A. hippocastanum and A. carnea with bleeding canker symptoms were examined. Subject of the screening were the pathogens Pseudomonas syringae pv. aesculi and Phytophthora spp. The trees were selected according to the macroscopically recorded damage pattern and the spatial distribution in the urban area. The sampling procedure was performed in a minimally invasive manner. The molecular biology examination was carried out on the day of sampling, allowing the laboratory findings to be available the following day. In total, 82 horse chestnut trees were tested positive for P. syringae pv. aesculi and 7 horse chestnut trees for Phytophthora spp. The sequencing revealed that the Phytophthora pathogen is Phytophthora cactorum. In addition, co-infection with both pathogens was found in 3 Horse Chestnut trees.
Key words: Pseudomonas syringae pv. aesculi, Phytophthora cactorum, Aesculus hippocastanum, Aesculus carnea, PCR, Hamburg, Bark disease
Pseudomonas syringae Bakterien kommen in mehr als 50 genetisch differenzierten Pathovarietäten an Gehölzen vor. Bereits in 26 Ländern wurden seit der Jahrtausendwende 55 Krankheitsausbrüche an Holzgewächsen dokumentiert, davon knapp die Hälfte in Europa (Lamichhane et al., 2014). Der wirtsspezifische Schaderreger Pseudomonas syringae pv. aesculi (Pae) wurde erstmals 2007 im deutschsprachigen Raum durch eine molekulare Identifizierung in Hamburg nachgewiesen (Dujesiefken et al., 2008). Charakteristische Anzeichen der Pseudomonas-Rindenkrankheit sind punktförmige, rostbraune Leckstellen am Stamm und in der Krone (ebd. 2008). Im Winter 2011/2012 wurden weitere Sekundärerreger an Rosskastanien mit Ausflusssymptomen entdeckt (Gaiser et al., 2013). Die Komplexerkrankung, an der unterschiedliche Basisdiomyceten beteiligt sind, folgt der bakteriellen Infektion mit dem Pathovar aesculi (Gaiser et al., 2013; Müller-Navarra et al., 2014), daher gilt dieser als Primärerreger (Dujesiefken, 2018). „Blutende“ Rosskastanien wurden bereits Mitte der 1970er Jahre in Großbritannien beschrieben – Horse Chestnut Bleeding Cancer. Der Auslöser war der eukaryotische Organismus Phytophthora (Brasier und Strouts, 1976). Geographisch sind Phytophthora-Arten weltweit verbreitet und befallen je nach Art, einen sehr engen oder weiten Wirtspflanzenkreis (Werres, 2011). In Deutschland haben Werres et al. (1995) Phytophthora-Arten an größeren Rosskastanienbeständen erstmals Mitte der 1990er Jahre als Krankheitserreger identifiziert. McEvoy et al. (2016) untersuchten 1587 Rosskastanien in Irland und fanden bei 61% der untersuchten Bäume Ausflusssymptome. Eine Stichprobenanzahl (n = 61) der Bäume wurde molekularbiologisch untersucht. Keine Probe enthielt Phytophthora Erreger, aber rund ein Drittel die DNA von P. syrinage pv. aesculi (ebd., 2016). Das Rosskastanien-Sterben im öffentlichen Grün wird auch in Deutschland mit P. syrinage pv. aesculi assoziiert (Dujesiefken und Gaiser, 2014). Die Gattung Aesculus ist ein fester Bestandteil im urbanen Stadtgrün der Freien und Hansestadt Hamburg. Rund 8400 Rosskastanien sind als Einzelbaum im digitalen Baumkataster erfasst, wovon ca. 6200 Exemplare an den Straßen stehen (Melzer, 2018). Bereits 2007 wurde die erste Reihenuntersuchung an 100 Rosskastanien mit Ausflusssymptomen im Stadtgebiet durchgeführt. Die Auswahl der Bäume erfolgte zufällig über eine Auswertung des digitalen Baumkatasters nach dem Kriterium „Ausfluss“. Die Phytopathogene P. syringae pv. aesculi und Phytophthora spp. wurden mikrobiologisch diagnostiziert. Eine eindeutige molekularbiologische Bestimmung des Pathovars aesculi war zu der Zeit nicht möglich. Um eine Verwechslung mit Pseudomonas syringae pv. syringae auszuschließen, wurden die 20 positiven Nachweise mittels der Polymerasekettenreaktion (PCR) molekularbiologisch untersucht (Hinz und Sadowska-Rybak, 2008). In 32 Proben konnten mittels traditioneller Kultivierung Phytophthora-Erreger nachgewiesen werden, die aber keiner Art zugeordnet wurden. Die 20 Pae-Verdachtsbäume wurden teilweise im Rahmen der zweiten Reihenuntersuchung 2014 erneut untersucht und der Verdacht bestätigt. Mit dem von Schmidt et al. (2009) entwickelten Schnelltest war die molekularbiologische Identifizierung über das Genom des Bakteriums möglich. Mit dem PCR-Verfahren wurde 2014 ausschließlich Pae diagnostiziert. Rund 40% der Bäume konnten der Pseudomonas-Rindenkrankheit zugeordnet werden (Dorendorf und Ludewig, 2014). In Ankara wurden an Rosskastanien mit Ausflusssymptomen und Absterbe-Erscheinungen in der Krone ausschließlich Phytophthora-Erreger diagnostiziert (Intini et al., 2002; Akıllı et al., 2012). Bei der ersten Reihenuntersuchung in Hamburg kamen Phytophthora-Erreger in 30% der untersuchten Proben vor. Eine Verwechslung der Pathogene kann aufgrund der ähnlichen Symptomatik daher nicht ausgeschlossen werden (Werres und Wagner, 2015; Dujesiefken et al., 2018). An vereinzelten Rosskastanien haben Gaiser et al. (2013) zudem beide Phytopathogene in Nordrhein-Westfalen nachgewiesen, eine Koinfektion ist daher ebenso denkbar. 2007 wurden in Hamburg bei der Probenentnahme Rindenfenster von ca. 10 bis 20 cm in der Höhe und 5 bis 10 cm in der Breite bis zum Splintholz reichend entnommen. 2014 war das entnommene Material zwar deutlich geringer, aber es entstanden erneut Wunden, die bis zum Splintholz reichten. Auch an nachweislich gesunden Bäumen blieben nach den Untersuchungen größere Verletzungen zurück, die als mögliche Eintrittspforten für P. syringae pv. aesculi oder andere Schaderreger fungieren könnten (Schmidt et al., 2014). Das Projektziel der dritten Reihenuntersuchung war die Differenzialdiagnose von P. syringae pv. aesculi und Phytophthora spp im öffentlichen Grün der Freien und Hansestadt Hamburg. Die Rosskastanien wurden nach den makroskopisch erfassten Symptomkomplexen und der räumlichen Verteilung im Stadtgebiet ausgewählt. Der mögliche Unsicherheitsfaktor während der visuellen Begutachtung, war daher ein zentrales Element dieser Reihenuntersuchung. Das Probenmaterial wurde molekularbiologisch über das PCR-Verfahren mit spezifischen Primerpaaren untersucht und eine Teilmenge sequenziert, um die Ergebnisse zu verifizieren und die Phytophthora-Art zu bestimmen.
Symptomatische Rosskastanien werden neben der regulären Bezirksbaumkontrolle und der Dokumentation im Baumkataster seit 2016 in einer gesonderten Geodatenbank in der Behörde für Umwelt und Energie (BUE) erfasst. Seit 2017 führt die BUE zudem ein umfangreiches Rosskastanien-Monitoring durch (Melzer und Doobe, 2019). Bisher sind in Hamburg ausschließlich die weißblühende Art A. hippocastanum und die rotblühende Art A. carnea betroffen. Aktuell stehen rund 600 Rosskastanien im Stadtgebiet unter Verdacht, mit Pae infiziert zu sein. (Melzer, 2018). Aus dieser Teilmenge wurden zwischen Juni und August 2018 94 Bäume beprobt. Die Schadsymptomatik der ausgewählten Verdachtsbäume deckt sich in der Regel mit den Symptomkomplexen, wie sie von Werres und Wagner (2015) oder Dujesiefken et al. (2018) für die beiden Pathogene detailliert beschrieben wurden. Vor der Probenentnahme erfolgte die Tauchdesinfektion der Arbeitsmaterialien in Natriumhypochlorid, zur chemischen Inaktivierung der Bakterien und des eukaryotischen, pilzähnlichen Organismus Phytophthora. Das am Arbeitsmaterial verbleibende organische Material wurde nach der Probenahme mittels physikalischem Desinfektionsverfahren thermisch entfernt (Bandte und Büttner, 2006; Kehr et al., 2010). Eine Probenmenge von ungefähr 300 mg wurde aus dem sekundären Phloem, im Übergangsbereich frischer Leckstellen, zwischen gesundem und nekrotischem Gewebe entnommen. Dazu wurde an einer möglichst kleinen, frischen Leckstelle die Borke entfernt, das Probenmaterial mit einem Stecheisen entnommen und mit einer Pinzette in den Probenbehälter (Sarstedt Schraubröhre 50 ml) überführt. Das Gefäß mit dem Probenmaterial wurde direkt gekühlt und noch am selben Tag in das Labor eingeliefert. Der stets zackige Verlauf im Übergangsbereich, die rötliche Färbung und die mosaikartige Scheckung sind für die Pseudomonas-Rindenkrankheit typisch (Abb. 1) (Dujesiefken et al., 2008; Kehr et al., 2010). Die für die Standard-PCR verwendeten Positiv-Kontrollen für Pseudomonas syringae pv. aesculi und Phytophthora spp. wurden vom Pflanzenschutzdienst Hamburg zur Verfügung gestellt. Als Negativkontrolle wurden Milli-Q Wasser und Gesamt-DNA aus einer nicht symptomatischen Rosskastanie verwendet.

Abb. 1. Nekrotisches Rindengewebe (dunkel) mit zackigem Übergangsbereich
Die Gesamt-DNA Extraktion erfolgte aus diesem Übergangsbereich, bei dem die Aktivität der Erreger am höchsten ist (Kehr et al., 2010; Werres, 2011). Das Probenmaterial wurde im Labor direkt aus dem Probenbehälter in einen Einmalbeutel (BioReba Extraction Bag) überführt. Unter Zugabe von 3–4 ml Extraktionspuffer (50 mM Na-PO4 pH 7,0) wurde das Material mit einem Hammer homogenisiert und die flüssige Phase des Homogenats in ein 1,5 ml Reaktionsgefäß pipettiert. Aus der Lösung wurden anschließend 100 μl für die DNA Extraktion verwendet. Die DNA Isolation mit dem DNeasy Plant Mini Kit (Qiagen) wurde nach Angaben des Herstellers durchgeführt. Die photometrische Bestimmung von Reinheit und Konzentration der zu untersuchenden Nukleinsäuren erfolgte mittels NanoDrop OneC (Thermo Scientific). Für die Ermittlung wurden je 2 μl Nukleinsäure-Lösung eingesetzt. Als Template für die Standard-PCR diente aus dem Probenmaterial extrahierte Gesamt-DNA. Der molekularbiologische Nachweis von Pseudomonas syringae pv. aesculi erfolgte durch Amplifikation der partiellen Gyrase B Sequenz mit den Primerpaaren G1/G2 (Schmidt et al., 2008) und G5/G6 (Schmidt et al., 2009). Dabei wurden 30 Zkylen von 30 s bei 94°C, 30 s bei 60°C und 30 s bei 72°C mit einer initialen Denaturierung von 2 min bei 94°C und einer finalen Elongation von 5 min bei 72°C durchgeführt. Der Nachweis einer Phytophthora spp. Infektion der Rosskastanien erfolgte mittels Standard-PCR mit dem Primerpaar YPh1F/YPh2R aus dem ras-bezogenen Proteingen (Exon 3 und Exon 6) entwickelt für die Detektion von ca. 15 Gehölz gefährdenden Phytophthora-Arten (Schena et al., 2007). Die PCR wurde nach dem gleichen Schema wie oben beschrieben durchgeführt. Dabei wurde lediglich das Annealing auf 58°C bei 45 s geändert und die finale Elongation um 5 min erhöht. Zur Erhöhung der DNA-Ausbeute auf Phytophthora spp. wurde dem Reaktionsansatz 5% DMSO (Farell and Alexandre, 2012) hinzugefügt und die Anzahl der Zyklen von 30 auf 40 erhöht. Nach der Gelelektrophoretischen Auftrennung (75 V für 45 min) wurden die PCR-Fragmente mittels UV-Transilluminator sichtbar gemacht und für die Sequenzierung mit dem innuPREP Gel Extraction Kit (Analytic Jena) extrahiert. Die PCR-Produkte wurden durch Eurofins Genomics (Deutschland) sequenziert. Die Auswertung der Sequenzen erfolgte mit den Programmen MEGA (Version 6.06, megasoftware) und BLAST (Altschul et al., 1997).
Insgesamt erfolgte bei 89 der 104 untersuchten Proben ein positiver Nachweis von Pseudomonas syringae pv. aesculi. Dabei wurde in allen 89 Proben das PCR-Fragment mit dem Primerpaar G5/G6 amplifiziert. Im Vergleich dazu zeigten nur 84 Proben einen positiven Nachweis mit dem Primerpaar G1/G2. Exemplarisch dafür sind die in Abb. 2 aufgeführten Proben B4 und M3.
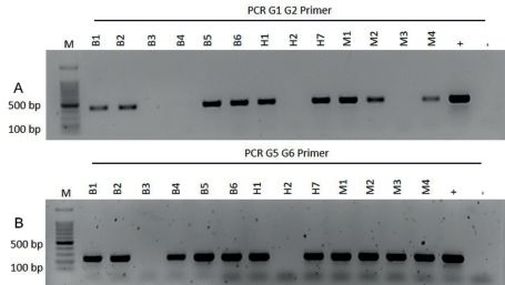
Abb. 2. Gelelektrophoretische Auftrennung von PCR-Produkten aus Phloem symptomatischer Aesculus. spp in 1%igem Agarosegel mit Pseudomonas syringae pv. aesculi spezifischem Primerpaar G1/G2 (A) und G5/G6 (B)
M = BenchTop 100 bp DNA Ladder, += Pseudomonas syringae pv. aesculi (Positivkontrolle), -= nicht symptomatische Kastanie (Negativkontrolle), B1, B2, B3, B4, B5, B6, H1, H2, H7, M2, M3, M4 = Proben zur Reihenuntersuchung
Die 104 untersuchten Proben verteilen sich auf 94 Rosskastanien. Davon gehören 74 Exemplare zu der weißblühenden Art A. hippocastanum und 20 zu der rotblühenden Art A. carnea. Abzüglich der Bäume, die mehrfach beprobt wurden, konnte bei 82 Einzelbäumen Pae nachgewiesen werden. Das Bakterium P. syringae pv. aesculi hat sich homogen in Hamburg verbreitet (Abb. 3). Die positiven Pae-Nachweise sind als roter Punkt auf der Karte dargestellt und die negativen als grüner. Die aktuellen Verdachtsbäume, die mit der Pseudomonas-Rindenkrankheit assoziiert werden, sind blau gekennzeichnet. Die weißen Punkte bilden den erfassten Einzelbaumbestand der Rosskastanien im Stadtgebiet ab.
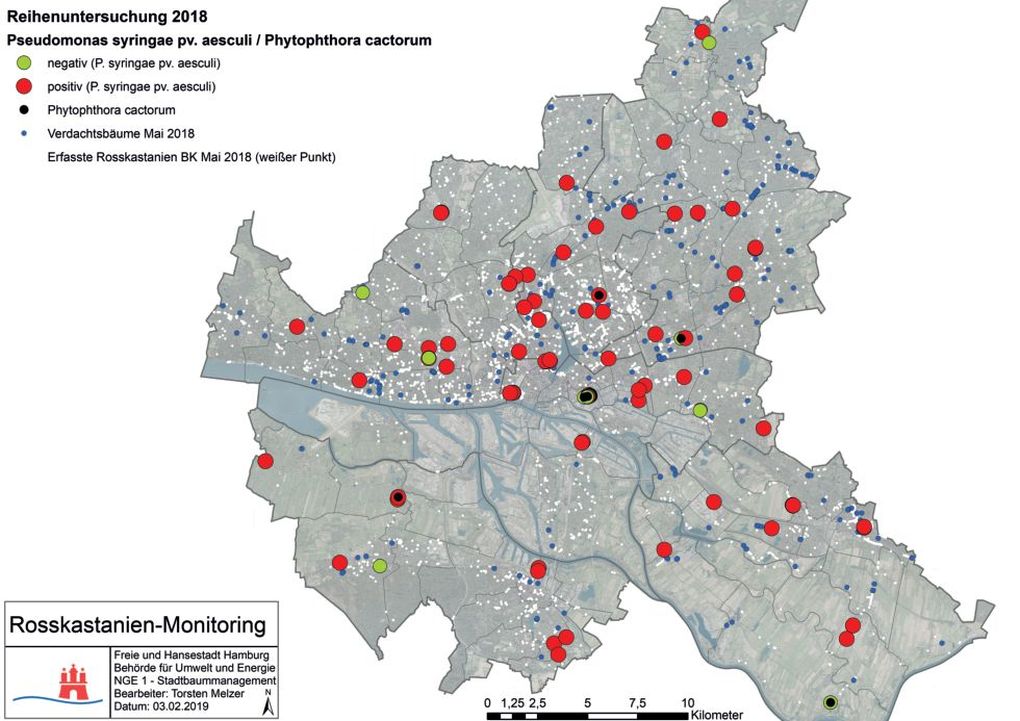
Abb. 3. Ergebnisse der Reihenuntersuchung 2018 in Hamburg
Ein Phytophthora-Befall konnte bei 7 Rosskastanien der Art A. hippocastanum, nachgewiesen werden. Diese Rosskastanien sind als schwarze Punkte auf der Karte dargestellt (Abb. 3). Die Sequenzierung ergab, dass es sich bei allen Proben um die Phytophthora-Art cactorum handelt. An 3 der 7 Rosskastanien wurde eine Koinfektion mit P. syringae pv. aesculi festgestellt. Die Koinfektion wurde jeweils aus dem gleichen Probenmaterial, sprich aus einer Probeentnahmestelle, diagnostiziert. An 8 Rosskastanien konnte keiner der untersuchten Schaderreger nachgewiesen werden. Zudem wurde ein großes Spektrum der in Hamburg vorkommenden Altersklassen abgedeckt. Die Altersklassen wurden in die Jugendphase (0–15 Jahre), in die Reifephase (15–80 Jahre) und in die Alterungsphase (> 80 Jahre) eingeteilt. Die Zählung der Jahre beginnt mit dem Pflanzjahr am Standort (Rust, 2013). Die Reifephase ist in die 1. Reifephase bis 40 Jahre am Standort und die 2. Reifephase bis 80 Jahre am Standort unterteilt. Die Rosskastanien mit einer Koinfektion wurden von der Gesamtsumme abgezogen (Tab. 1).
Tab. 1. Laborbefund der 94 untersuchten Rosskastanien in Altersklassen unterteilt
| Jugendphase | 1. Reifephase | 2. Reifephase | Alterungsphase | Gesamt |
(+) Pae | 6 | 43 | 17 | 16 | 82 |
(+) P. cactorum | – | 5 | – | 2 | 7 |
(+) Negativ | – | 4 | 4 | – | 8 |
(-) Koinfektion | – | 2 | 0 | 1 | 3 |
(=) Gesamt | 6 | 50 | 21 | 17 | 94 |
Neben den vielen klaren Befunden während dieser Reihenuntersuchung kam es auch zu unklaren Ergebnissen. Die Beprobung älterer Leckstellen führte in der ersten Hälfte des Projektzeitraums zu zwei negativen Befunden, die bei einer weiteren Probenentnahme widerlegt wurden. Daher wurde an 4 Rosskastanien, mit frischen und eingetrockneten Leckstellen, eine doppelte Beprobung durchgeführt (n = 8). Bei allen acht Proben wurde P. syringae pv. aesculi nachgewiesen. Eine Rosskastanie im Bezirk Harburg zeigte typische Leckstellen der Pseudomonas-Rindenkrankheit im oberen Stammbereich, aber am Stammfuß eine Leckstelle, die auf einen Phytophthora-Befall schließen ließ. Die Probe am Stammfuß enthielt beide Schaderreger – die Probe am oberen Stamm ausschließlich den Pathovar aesculi. Eine Rosskastanie in Hamburg Nord mit sehr starkem Ausfluss am Stammfuß, Stamm und im Kronenbereich wurde, um eine Koinfektion auszuschließen, an drei Stellen beprobt. In allen drei Proben wurde ausschließlich das Bakterium P. syringae pv. aesculi nachgewiesen. An einem Altbaum in Hamburg Bergedorf mit ähnlichem Schadbild, allerdings weiter fortgeschritten, wurden ebenfalls zwei Proben entnommen. Eine am Stammfuß und eine im oberen Stammbereich. Bei dieser Entnahme wurde ausschließlich der Erreger Phytophthora cactorum nachgewiesen. Zudem wurde von zwei im Februar gefällten Altbäumen – der Stammfuß war noch nicht gerodet – aus frischen Leckstellen Probenmaterial entnommen. Der Pathovar aesculi konnte auch hier nachgewiesen werden. Der Nachweis und die Verifizierung der Ergebnisse erfolgten mittels Blast-Analyse (BlastN) (Altschul et al., 1997). Dazu wurden zwei der Proben, in denen jeweils nur mittels G5/G6 Primerpaar ein Fragment generiert werden konnte, sequenziert.
Dabei zeigten die als P. syringae pv. aesculi positiv deklarierten Proben eine Sequenzübereinstimmung von 100% zur partiellen Gyrase B Sequenz von Pseudomonas syringae pv. aesculi. Alle mit den Primern YPh1F/2R amplifizierten Fragmente wurden sequenziert und stimmten zu 100% mit Phytophthora cactorum überein.
Die Reihenuntersuchung im Sommer 2018 war bereits die dritte in Hamburg (Tab. 2). Da die Methodik der Untersuchung sich in den Jahrgängen unterscheidet, sind die Ergebnisse nur bedingt vergleichbar. Dennoch zeigen sie, dass seit 2007 Hamburgs Rosskastanien von beiden Schaderregern befallen werden und die seither gewonnenen Erkenntnisse dazu geführt haben, dass die visuelle Einschätzung der Symptome deutlich höher mit dem Laborbefund korreliert. Hervorzuhebende Unterschiede bei der methodischen Herangehensweise zwischen 2007 und 2018 waren: die zufällige und gezielte Baumauswahl, die mikrobiologische und molekularbiologische Laboranalytik sowie die Vorgehensweise während der Probenentnahme. In den Untersuchungen von 2014 und 2018 wurde nach der von Schmidt et al. (2009) entwickelten Methodik vorgegangen, um eine Vergleichbarkeit der molekularbiologischen Untersuchung zu gewährleisten.
Tab. 2. Diagnostizierte Pathogene der Reihenuntersuchungen zwischen 2007 und 2018 in Hamburg
Jahr; Anzahl/Bäume | 2007 (n = 93) | 2014 (n = 101) | 2018 (n = 94) |
P. syringae pv. aeculi | 20 (P. syringae) | 43 (Pae) | 82 (Pae) |
Phytophthora spp. | 33 (lediglich die Gattung bestimmt) | nicht untersucht | 7 (P.cactorum) |
Koinfektion | 7 (Phy.spp/P.syringae) | entfällt | 3 (P.cactorum/Pae) |
Probengesamtzahl | 93 | 101 | 94 |
Die räumliche Verteilung zeigt deutlich, dass in fast allen Stadtteilen, in denen Rosskastanien stehen, nachweislich das Bakterium vorkommt. Der makroskopische Verdacht der MitarbeiterInnen aus der Hamburger Baumkontrolle wurde in ca. 90% aller Fälle bestätigt. Dies zeigt, dass in Hamburg bereits eine sehr sichere visuelle Einschätzung zum Pathogenverdacht in der Baumkontrolle besteht. Im Abschlussbericht der zweiten Reihenuntersuchung 2014 wurde auf eine erneute Beprobung unterschiedlicher Standorte hingewiesen. Der Probeentnahmezeitpunkt war bereits im Oktober und die Leckstellen waren eingetrocknet. Der Befund lag an der Nachweisgrenze oder das negative Laborergebnis passte nicht zu dem dokumentierten Symptomkomplex (Dorendorf und Ludewig, 2014). Ein Grund hierfür könnte die Beprobung sehr alter Leckstellen außerhalb des Übergangsbereiches sein. Die Probeentnahme an bereits eingetrockneten Leckstellen (n = 2) führte zu Beginn dieser Reihenuntersuchung ebenfalls zu unklaren Ergebnissen. Daher stellte sich die Frage, wie hoch der Unsicherheitsfaktor bei der Beprobung älterer Leckstellen ist. Versuchsweise wurde deshalb an vier Rosskastanien mit eingetrockneten und frischen Leckstellen (n = 8) eine doppelte Probenentnahme durchgeführt. Entgegen der erwarteten Annahme, dass eine Teilmenge der Proben einen Negativnachweis erhält, wurde bei jeder Probe eindeutig das Bakterium P. Syringae pv. aesculi nachgewiesen. Die anfänglich angenommene Hypothese, das eingetrocknete alte Leckstellen zu einem negativen Ergebnis führen müssen, wurde dadurch falsifiziert. Die Untersuchungen dieser Studie bestätigen demnach nur, mit einen kleinen Stichprobenumfang, dass die Auswahl der Leckstelle ein Unsicherheitsfaktor für das Ergebnis der Analyse sein kann. Im Weiteren wird das abgestorbene Rindengewebe nach dem Gewebetod durch Sekundärerreger besiedelt (Kehr et al., 2010). Eine konsequente Desinfektion der Werkzeuge und die Probenentnahme an frischen Leckstellen, im Übergangsbereich des nekrotischen Rindengewebes sind ausschlaggebend für zuverlässige und replizierbare Ergebnisse. Kehr et al. (2010) weisen darauf hin, dass Ethanol die DNA der Pathogene nicht zerstört und damit das sensitive PCR-Verfahren bei mangelnder Desinfektion zu fehlerhaften Ergebnissen führt. Dies ist bei der DNA-Extrahierung im Labor und bei der Probeentnahme zu beachten, um eine Verschleppung zu vermeiden (Kehr et al., 2010; Fröhlich et al., 2016b). Die Probenaufbereitung wurde daher in einem Einmalbeutel durchgeführt. Dieses Vorgehen hat sich bei der großen Probenstückzahl bewährt und das Risiko einer Verschleppung bei der Zerkleinerung im Labor konnte dadurch deutlich minimiert werden. Die Übergangsstelle sollte daher bei der Probenahme identifiziert und eine ausreichende Materialmenge aus dem sekundären Phloem entnommen werden. Die Borke und der Schleimfluss müssen entfernt werden. Den Ausfluss besiedeln zahlreiche saprophytische Bakterien und Pilze (Kehr et al., 2010). Als besonders geeignet haben sich kleine Leckstellen erwiesen, da in diesem Bereich nur wenig Borke entfernt werden muss. Abb. 4 zeigt eine solche Leckstelle, dieselbe Leckstelle in 20-facher Vergrößerung und das darunter liegende nekrotische Rindengewebe.

Abb. 4. Leckstelle, 20-fach vergrößert und Phloem-Nekrose
Um eine möglichst baumschonende, minimalst invasive Probenentnahme durchzuführen, wurde in der Testphase mit Kleinstmengen experimentiert. Ein Probenstück von ca. 300 mg ist für den routinemäßigen Nachweis von Pseudomonas syringae pv. aesculi und Phytophthora spp. geeignet (Kehr et al., 2010). Allerdings sollten die Proben den charakteristischen Grenzbereich zwischen nekrotischem und nicht befallenem Gewebe beinhalten, der die höchste Aktivität der Pathogene aufweist (Green et al., 2009; Kehr et al., 2010; Werres, 2011). In der Regel wurde eine Probe pro Baum entnommen. Weitere Proben am selben Baum wurden nur bei unklarem Ergebnis oder bei einer Unsicherheit während der Baumansprache genommen. In den vorangegangenen Reihenuntersuchungen in Hamburg wurde noch bis zum äußeren Splintholz Rindengewebe entnommen. Kehr et al. (2010) empfehlen ebenfalls eine Probentiefe bis zum äußeren Splintholz. Für die molekularbiologische Analyse wurden in dieser Untersuchung Kleinstmengen aus dem Bast entnommen. Die geringe Menge an Probematerial reduziert den Materialaufwand für die Probenahme, erleichtert die Weiterverarbeitung im Labor und bewirkt durch kleinere und flachere Wunden geringeren Aufwand an Abwehrreaktionen des zu beprobenden Baumes (Shigo, 1984). Das Rindengewebe verfügt über unterschiedliche Strategien zur Wundheilung. Das Bast-Gewebe verfärbt sich nach dem Eingriff dunkel und die Ausdehnung nach der Verletzung ist aufgrund des höheren Parenchymanteils und der kleineren Siebröhren deutlich geringer als im tieferliegenden Gewebe. Der beschädigte Bereich wird linsenförmig abgeschottet und die Parenchymzellen teilen sich parallel zum Wundrand und bilden das Wundperiderm (Dujesiefken et al., 2008). Die natürliche Wundreaktion wird unterstützt und Eintrittspforten für weitere Pathogene werden schneller verschlossen. Die natürliche Wundreaktion der beprobten Bäume zeigen (Abb. 5) und (Abb. 6). Die Bilder entstanden während einer der ersten Probenentnahmen am 03.07.2018 und ca. ein halbes Jahr später am 11.01.2019 bei einer erneuten Begutachtung des Baumes.

Abb. 5. Probenentnahmen am 03.07.2018

Abb. 6. Wundreaktion: ca. 6 Monate später
Die Probenahmestelle von 2007 wurde bereits mit einem Flächenkallus überwallt, der Eingriff 2014 war bereits deutlich kleiner, aber reicht auch bis zum Splintholz. Das Rindenfenster wurde nicht überwallt. 2018 wurde das Probenmaterial aus dem sekundären Phloem des Bast-Gewebes entnommen (Abb. 7). Die Rosskastanie wurde bei jeder Analyse positiv auf P. syringae pv. aesculi getestet.

Abb. 7. Entwicklung der Probenentnahme von 2007 bis 2018 in Hamburg
Mitunter reagiert die Gattung Aesculus aus unterschiedlichsten Gründen mit Ausfluss, diese können ebenso durch Verletzungen hervorgerufen oder auch gänzlich unspezifischer Natur sein (Dujesiefken et al., 2018). Auf dem Billstedter Marktplatz, in Hamburg Mitte, wurden zwei Rosskastanien beprobt die absterbende Kronenpartien und Ausflusssymptome zeigten. Die beiden Proben waren negativ und ein möglicher Grund für die angetroffene Schädigung der Bäume könnte auf die städtischen Stressfaktoren zurückzuführen sein. Rust (2013) beschreibt die vitalitätsgefährdende Verkettung unterschiedlicher Stressfaktoren, die in der Stadt vorkommen und zu nachhaltigen Baumschäden führen. Am Billstedter Marktplatz sind dies der mutmaßliche Streusalzeinsatz, die starke Bodenverdichtung, der hohe Versiegelungsgrad und der regelmäßig stattfindende Wochenmarkt mit den Auf- und Abbauarbeiten. Der molekularbiologischen Untersuchung sollte daher eine detaillierte Begutachtung und Dokumentation am Standort vorangehen (Werres, 2011). Die Baumansprache und Begutachtung der drei Symptomkomplexe – Phytophthora-Erkrankung, Pseudomoas-Rindenkrankheit und Komplexkrankheit der Rosskastanie – wurde von Werres und Wagner (2015) oder Dujesiefken et al. (2018) detailliert beschrieben. Bei unklarer Symptomatik oder bei Problemen während der Probenahme, sollte mindestens eine weitere Probe am Einzelbaum entnommen werden (Kehr et al., 2010). Die Phytophthora-Art cactorum ist weltweit verbreitet und hat einen ungefähren Wirtspflanzenkreis von 1560 Pflanzengattungen in 54 Familien (Werres, 2011). In Deutschland wies Jung (2009) P. cactorum als häufigstes Pathogen der Gattung Phytophthora in urbanen Baumbeständen an der Gattung Fagus nach. Für die Gattung Aesculus kommen die Hamburger Untersuchungen zu dem gleichen Schluss. Cerny et al. (2009) haben in Tschechien an Fagus sylvatica und Aesculus hippocastanum ebenfalls P. cactorum als Schaderreger diagnostiziert. In einer Parkanlage in Ankara wurde 2002 erstmalig in der Türkei Phytophthora cactorum nachgewiesen. Ausflusssymptome und absterbende Teile in der Krone wurden auch hier an Rosskastanien im Zusammenhang mit P. cactorum dokumentiert (Intini et al., 2002). Im Jahr 2012 wurde in Ankara ein weiterer Phytophthora-Erreger festgestellt, aber an keiner der untersuchten Rosskastanien mit Ausflusssymptomen wurde Pae diagnostiziert (Akıllı et al., 2012). Im westlichen Europa wurden die ersten Fälle von Pae aus den Niederlanden und Großbritannien gemeldet (Dujesiefken et al., 2008). Den östlich gelegensten empirischen Nachweis haben Pirc et al. (2018) in Slowenien erbracht. Daher lässt sich vermuten, dass Pae noch nicht von Westeuropa bis zum Bosporus vorgedrungen ist, aber ein ostwärts Trend ist zu beobachten. In Irland erfolgte der Erstnachweis von Pae bereits 2010 (EPPO, 2018). McEvoy et al. (2016) konnten an keiner Rosskastanie mit Ausflusssymptomen Phytophthora-Erreger feststellen. Obwohl Phytophthora-Erreger in Irland seit dem 19. Jahrhundert erfasst wurden und inzwischen ca. 27 unterschiedliche Arten auf der Insel diagnostiziert wurden (O’Hanlon et al., 2016). Bei der stichprobenartigen Untersuchung wurde neben weiteren Erregern hauptsächlich P. syrinage pv. aesculi mit dem Verfahren der Realtime-PCR ermittelt (McEvoy et al., 2016). Die Studien aus Irland und der Türkei lassen Rückschlüsse auf Hamburg zu. In der Freien und Hansestadt ist die Phytophthora-Erkrankung, ähnlich wie in Ankara, zwar vorhanden, aber es sind keine größeren Bestände betroffen, sondern nur Einzelbäume. 61% der 1537 untersuchen Rosskastanien in Irland zeigten Ausflusssymptome. Die Untersuchung einer Stichprobe von 61 Bäumen ergab, dass 20% der untersuchten Proben die DNA der Pae Bakterien enthielten (ebd., 2016). Rund 460 Rosskastanien mit Symptomen der Komplexkrankheit mussten seit 2013 in Hamburg gefällt werden und rund 600 Rosskastanien zeigen aktuell Symptome der bakteriellen Infektion. Es sind nicht nur Einzelbäume, sondern ganze Straßenzüge betroffen. Zusammengerechnet sind dies rund 15% der erfassten Rosskastanien – ausgehend vom Bestand 2013 (Melzer, 2018). Dies zeigt, dass Pae an der Rosskastanie deutlich öfter vorkommt und die Verbreitung wesentlich schneller ist. P. syringae pv aeculi gilt als Primärerreger der Komplexerkrankung (Dujesiefken, 2018) und die Ergebnisse dieser Reihenuntersuchung bestätigen Pae als verantwortliches Phytopathogen für das Rosskastanien-Sterben im öffentlichen Grün der Freien und Hansestadt Hamburg. Durch den molekularbiologischen Schnelltest nach Schmidt et al. (2009) standen bereits am Folgetag die Ergebnisse zur Verfügung, konnten ausgewertet und mit den aufgenommenen Standortdaten verglichen werden. Eine erneute Probenahme erfolgte bei unklarem Ergebnis. Ein möglicher Einflussfaktor für die geringere Zuverlässigkeit der G1/G2-Primer könnte die Länge der PCR-Fragmente in Zusammenhang mit dem jeweiligen Pathogen-Titer der untersuchten Probe spielen. Probleme im Nachweis größerer PCR-Fragmente aus Gehölzen konnte bereits bei dem prunus necrotic ringspot virus (PNRSV) nachgewiesen werden (Rosner et al., 1997). Dabei konnte gezeigt werden, dass Primer, die ein kurzes PCR-Fragment amplifizieren, sowohl höhere Mengen an PCR-Produkt generieren, als auch bei geringem Pathogen-Titer einen zuverlässigeren Nachweis erbrachten. Da sich alle Gyrase B Primer zur routinemäßigen Untersuchung von Pseudomonas syringae pv. aesculi an Aesculus spp. eignen (Schmidt et al., 2009), sollte unter Berücksichtigung der in diesem Projekt beobachteten Unterschiede in der Sensitivität vorrangig das Primerpaar G5/G6 für den Pathogen-Nachweis verwendet werden. Die Quantifizierung der extrahierten DNA mittels NanoDrop zeigte sehr geringe DNA-Ausbeuten von 0,4 bis 13,9 ng/μl. Sowohl mit der geringsten als auch der höchsten DNA-Menge erfolgte ein positiver Nachweis für Pseudomonas syringae pv. aesculi mit beiden verwendeten Primerpaaren (G1/G2 und G5/G6). Weiterhin konnte keine Korrelation zwischen der Quantität der extrahierten DNA und dem positiven Nachweis von Pseudomonas syringae pv. aesculi oder Phytophthora spp. gefunden werden. Die beiden amplifizierten Pseudomonas-Fragmente zeigten eine eindeutige Übereinstimmung mit dem Pathogen Pseudomonas syringae pv. aesculi. Die Fragmente, amplifiziert mit den YPh1F/2R-Primern, zeigten eine eindeutige Homologie mit Phytophthora cactorum. Besonders beim Nachweis einer Koinfektion von Pseudomonas und Phytophthora sollte ein Nachweis mit erhöhter Sensitivität gewählt werden. So waren bei einem Nachweis von Pae die Banden, des amplifizierten PCR-Fragmentes, des Phytophthora-Erregers deutlich schwächer als in den weiteren positiven Phytophthora-Befunden (Abb. 8). Auffällig ist, dass mit Ausnahme der Probe W1 alle weiteren positiven Nachweise von P. cactorum nur nach Modifikation des PCR-Protokolls erfolgten. Um auswertbare PCR-Fragmente zu generieren, wurde daher das Additiv DMSO verwendet, das die Formation von Sekundärstrukturen in GC-reichen DNA-Templates unterbindet (Farell and Alexandre, 2012).
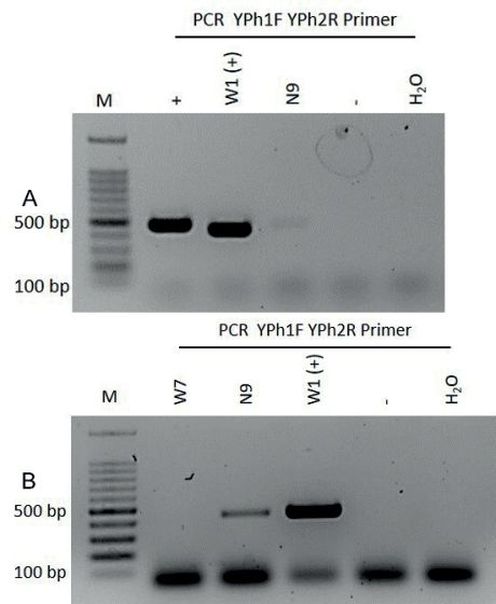
Abb. 8. Gelelektrophoretische Auftrennung von PCR-Produkten aus Phloem symptomatischer A. hippocastanum in 1%igem Agarosegel mit Phytophthora spp. spezifischem Primerpaar YPh1F/YPh2R
A: PCR-Ansatz ohne DMSO, PCR 30 Zyklen
B: PCR-Ansatz mit DMSO, PCR 40 Zyklen
M = BenchTop 100 bp DNA Ladder, += Phytophthora ramorum (Positivkontrolle), + (1:10)= Phytophthora ramorum und nicht symptomatische Kastanie im Verhältnis 1:10, -= nicht symptomatische Kastanie (Negativkontrolle), H2O = Milli-Q (Negativkontrolle), W1, W7, N9 = Proben zur Reihenuntersuchung
Neben einem allgemein geringen Pathogen-Titer kann zusätzlich die Präsenz von Pseudomonas einen destruktiven Einfluss auf Phytophthora ausüben. So konnte beispielsweise nachgewiesen werden, dass volatile organische Verbindungen, welche von verschiedenen Pseudomonaden abgesondert werden, eine inhibierende Wirkung auf das Wachstum von Phytophthora infestans haben (Hunziker et al., 2015). Dieser Effekt könnte eine Erklärung dafür sein, warum Phytophthora-Erreger nur an einer sehr geringen Anzahl von Rosskastanien identifiziert wurden. Gaiser et al. (2013) haben ebenfalls eine Koinfektion der Pathogene an Rosskastanien festgestellt. Bei der Untersuchung von 36 Rosskastanien mit Symptomen der Komplexkrankheit – verteilt auf drei Bundesländer – wurde an zwei Rosskastanien Pae und P. cactorum diagnostiziert. In den 104 untersuchten Proben dieser Reihenuntersuchung wurde an drei Rosskastanien mit Ausflusssymptomen eine Koninfektion mit Pae und P. cactorum festgestellt. Die Ergebnisse dieser Untersuchung und die von Gaiser et al. (2013) zeigen, dass Koinfektionen nicht auszuschließen sind. Dies sollte bei der Baumansprache, insbesondere bei Unsicherheiten zum makroskopisch erfassten Verdacht, berücksichtig werden.
In Hamburg müssen jährlich Rosskastanien mit Symptomen der Komplexkrankheit gefällt werden, weil die Verkehrssicherheit nicht mehr gewährleistet werden kann (Melzer und Doobe, 2019). Das Bakterium Pseudomonas syringae pv. aesculi gilt als Primärerreger der Komplexkrankheit der Rosskastanie (Dujesiefken, 2018) und wird als relativ junger Schaderreger in Europa mit dem Rosskastanienschäden im öffentlichen Raum assoziiert. (Dujesiefken und Gaiser, 2014; Werres und Wagner, 2015, McEvoy et al., 2016; Hauser, 2018; Dujesiefken, 2018). Langjährige Erfahrung in der visuellen Erfassung zu den Symptomkomplexen führt zu einer zuverlässigen Diagnose und ein flächendeckender Befall konnte im Stadtgebiet der Freien und Hansestadt Hamburg nachgewiesen werden. Die Differentialdiagnose der Pathogene Phytophthora spp. und Pseudomonas syringae pv. aesculi ist über das gleiche Probenahme-Verfahren mit anschließender molekularbiologischer Untersuchung zu empfehlen. Dabei sollte unbedingt der Übergangsbereich des nekrotischen Rindengewebes gewählt werden, da die Aktivität der Schaderreger hier am höchsten ist (Kehr et al., 2010; Werres, 2011). Das PCR-Verfahren ist sehr sensitiv und verbleibende Kleinstmengen am Werkzeug können zu einer Verschleppung der Schaderreger führen (Kehr et al., 2010, Fröhlich et al., 2016b). Die beschriebene Technik der Probenahme und die Vorgehensweise im Labor hat sich in diesem Projekt (n = 104) bewährt. Für die chemische Inaktivierung der Pathogene sollte ein Mittel mit Chlor, Chlordioxid oder Wasserstoffperoxid verwendet werden bzw. eine thermische Desinfektion erfolgen (Bandte und Büttner, 2006; Kehr et al., 2010). Die Baumansprache sowie eine detaillierte Begutachtung und Dokumentation der Krankheitssymptome sollten der molekularbiologischen Untersuchung vorangehen. Etwa 20% der zufällig ausgewählten Rosskastanien im Jahr 2007, konnten dem Verdacht der bakteriellen Infektion zugeordnet werden. 2014 waren es etwas mehr als 40% und 2018 fast 90%. Die hohe Steigerung ist nicht auf ein exponentielles Ansteigen der erkrankten Bäume zurückzuführen, sondern auf die bessere Einschätzung der Symptomkomplexe, die gezielte Probenentnahmetechnik, die verbesserte Labordiagnostik und die Dokumentation von Pae im Hamburger Baumkataster. Dennoch ist der aktuelle Anstieg besorgniserregend und ein Ausfall von weiteren Rosskastanien in den kommenden Jahren zu erwarten. Die folgende Komplexerkrankung der Rosskastanie ist in der Regel der Grund für Fällungen und die Symptomatik der bakteriellen Infektion führt nicht zwingend zu einem verkehrsgefährdenden Zustand der Bäume. Zumindest solange nicht, wie der Baum vital ist und keine weiteren Schaderreger auftreten (Dujesiefken et al., 2008). Ist ein Baum erkrankt, sollte dieser im Rahmen der Baumkontrollen halbjährlich kontrolliert werden, um die Vitalität während Vegetationsperiode und mögliche Pilzfruchtkörper in den Wintermonaten zu erkennen (Dujesiefken, 2018). In Hamburg werden Rosskastanien erst dann gefällt, wenn die Verkehrssicherheit nicht mehr gegeben ist. Negative Labornachweise von ersichtlich kranken Rosskastanien können mit der Qualität bzw. dem Alter der Leckstelle und einer zu geringen Aktivität der Erreger zusammenhängen. Bei Unsicherheiten sollte eine erneute Probenentnahme an einer frischen Leckstelle zu einem anderen Zeitpunkt durchgeführt werden. Für die molekularbiologische Untersuchung von Pae konnte mit größerer Sicherheit ein Fragment der Pae-DNA generiert werden, wodurch dementsprechend bei geringem Budget das G5/G6 Primerpaar die bessere Wahl ist. Die Zusammenarbeit zwischen dem Pflanzenschutzamt der Behörde für Wirtschaft, Verkehr und Innovation (BWVI) und dem Stadtbaummanagement der Behörde für Umwelt und Energie (BUE) wird im Jahr 2019 fortgesetzt. Das methodische Vorgehen dieser Reihenuntersuchung soll über den gesamten Jahresverlauf ausgeweitet werden und zum Erkenntnisgewinn der Aktivität von P. syringae pv. aesculi beitragen. Die Phytophthora-Erkrankung und die Verbreitung im Stadtgebiet werden weiter beobachtet, stehen aber aktuell nicht im Vordergrund. Die Diagnostik erfolgt daher in erster Linie nur noch bei einem makroskopischen Verdacht zum Symptomkomplex oder bei einem erhöhten Unsicherheitsfaktor während der Baumansprache.
Akıllı, S., Ç. Ulubas Serçe, Y. Katırcıoğlu, S. Maden, 2012: Phytophthora citrophthora, a new pathogen causing decline on horse chestnut in Turkey. Forest Pathology 42 (4), 299–304, DOI: 10.1111/j.1439-0329.2011.00754.x.
Altschul, S.F.T.L. Madden, A.A. Schäffer, J. Zhang, Z. Zhang, W. Miller, D.J. Lipman, 1997: Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Research 25 (17), 3389–3402, DOI: 10.1093/nar/25.17.3389.
Brasier, C.M., R.G. Strouts, 1976: New records of Phytophthora on trees in Britain. Forest Pathology 6 (3), 129–136, DOI: 10.1111/j.1439-0329.1976.tb00517.x.
Bandte, M., C., Büttner, 2006: Desinfektion zur Dekontamination von Pflanazenpathogenen Viren. In: Dujesiefken, D. (Hrsg.), Jahrbuch der Baumpflege 2013: Yearbook of Arboriculture. Braunschweig, Haymarket Media. 230-235.
Cerny, K., V. Strnadova, B. Gregorova, V. Holub, M. Tomsovsky, M. Mrazkova, S. Gabrielova, 2009: Phytophthora cactorum causing bleeding canker of common beech, horse chestnut, and white poplar in the Czech Republic. Plant Pathology 58 (2), 394, DOI: 10.1111/j.1365-3059.2008.01970.x.
Dorendorf, J., K. Ludewig, 2014: Voruntersuchung zu Ursachen für das Kastanienbaumsterben im öffentlichen Grün in Hamburg. Projekt Hamburg BUE/BWVI Pflanzenschutzdienst Archiv.
Dujesiefken, D., O., Schmidt, R., Kehr, H., Stobbe, U., Moreth, T.H., Schroeder, 2008: Pseudomonas-Rindenkrankheit der Rosskastanie – Erstnachweis des Bakteriums Pseudomonas syringae pv. Aesculi in Deutschland. In: Dujesiefken, D. (Hrsg.), Jahrbuch der Baumpflege 2018: Yearbook of Arboriculture. Braunschweig, Haymarket Media.153-164.
Dujesiefken, D., W. Liese, 2012: Das CODIT-Prinzip: Von den Bäumen lernen für eine fachgerechte Baumpflege. Braunschweig, Haymarket Media. 160 S.
Dujesiefken, D., O. Gaiser, 2014: Pseudomonas: Auslöser für das Rosskastanien-Sterben. AFZ Der Wald. Bd. 24. S. 35-38.
Dujesiefken, D., 2018: Aktuelles zur Pseudomonas-Rindenkrankheit und zum Rosskastanien-Sterben. Pro Baum 2018 (2), 10-15.
Dujesiefken, D., P., Jaskula, T., Kowohl, A., Lichtenauer, 2018: Baumkontrolle unter Berücksichtigung der Baumart: Bildatlas der typischen Schadsymptome und Auffälligkeiten. Braunschweig, Haymarket Media, 263-290.
EPPO, 2018: EPPO GLOBAL DATABASE, Distribution details in Ireland. Zugriff: 10 Dezember 2018, URL: https://gd.eppo.int/taxon/PSDMAX/distribution/IE.
Farell, E.M., G. Alexandre, 2012: Bovine serum albumin further enhances the effects of organic solvents on increased yield of polymerase chain reaction of GC-rich templates. BMC research notes 5, 257, DOI: 10.1186/1756-0500-5-257.
Fröhlich, H., K., Morgenstern, D., Krabel, A. Roloff, 2016: Pseudomonas in Dresden entdeckt. TASPO Baumzeitung 01/2016, 39-42.
Fröhlich, H., K., Morgenstern, D., Krabel, A. Roloff, 2016b: Pseudomonas-Nachweis in Dresden. AFZ der Wald, 4/2016, 34-37.
Gaiser, O., D. Dujesiefken, R. Kehr, 2013: Das neue Schadbild an der Rosskastanie – Untersuchungsergebnisse und Hinweise für die Praxis. In: Dujesiefken, D. (Hrsg.), Jahrbuch der Baumpflege 2013: Yearbook of Arboriculture. Braunschweig, Haymarket Media.229-237.
Green, S., B. Laue, C.G. Fossdal, S.W. A’Hara, J.E. Cottrell, 2009: Infection of horse chestnut (Aesculus hippocastanum) by Pseudomonas syringae pv. aesculi and its detection by quantitative real-time PCR. Plant Pathology 58 (4), 731–744, DOI: 10.1111/j.1365-3059.2009.02065.x.
Hauser, S., 2018: Ist Europas beliebteste Baumart noch zu retten? Ergebnisse eines Workshops zur Rosskastanie. Historische Gärten 2018, (1), 4-7.
Hunziker, L., D. Bönisch, U. Groenhagen, A. Bailly, S. Schulz, L. Weisskopf, 2015: Pseudomonas strains naturally associated with potato plants produce volatiles with high potential for inhibition of Phytophthora infestans. Applied and environmental microbiology 81 (3), 821–830, DOI: 10.1128/AEM.02999-14.
Hinz, Y., M. Sadowska-Rybak, 2008: Vorversuche zur Indentifizierung von phytopathogenen Erregern der Blutungskrankheit an Rosskastanien (Aesculus Hippocastanum) in Hamburg. Nachrichtenbl. Deut. Pflanzenschutzd., 60, 91-91.
Intini, M., M. Gurer, S. Ozturk, 2002: First Report of Bleeding Canker Caused by Phytophthora cactorum on Horse Chestnut in Turkey. Plant Disease 86 (6), 697, DOI: 10.1094/PDIS.2002.86.6.697C.
Jung, T., 2009: Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. Forest Pathology 39 (2), 73–94, DOI: 10.1111/j.1439-0329.2008.00566.x.
Kehr, R., P., Möhlenhoff, K., Petersen, 2010: Pseudomonas-Rindenkrankheit der Rosskastanie – Anleitung zur Probenahme für den Schnellnachweis. In: Dujesiefken, D. (Hrsg.), Jahrbuch der Baumpflege 2010: Yearbook of Arboriculture. Braunschweig, Haymarket Media.306-310.
Lamichhane, J.R., L. Varvaro, L. Parisi, J.-M. Audergon, C.E. Morris, 2014: Disease and Frost Damage of Woody Plants Caused by Pseudomonas syringae. Advances in Agronomy 126, 235-295, DOI: 10.1016/B978-0-12-800132-5.00004-3.
McEvoy, A., F. O'Regan, C.C. Fleming, B.P. Moreland, J.A. Pollock, B.W. McGuinness, T.R. Hodkinson, 2016: Bleeding canker of horse chestnut (Aesculus hippocastanum) in Ireland: incidence, severity and characterization using DNA sequences and real-time PCR. Plant Pathology 65 (9), 1419–1429, DOI: 10.1111/ppa.12529.
Melzer, T., 2018: Die Komplexkrankheit der Rosskastanie – Bestandsaufnahme und Verbreitungsdynamik auf Hamburger Stadtgebiet. In: Bouillon, J.M. (Hrsg.), 36. Osnabrücker Baumpflegetage: 4. und 5. September 2018: Tagung, Fachmesse, Workshops, Berlin, Hannover, Patzer Verlag, S.75-88.
Melzer, T., G. Doobe, 2019: Zustand der Hamburger Rosskastanien. TASPO Baumzeitung 1, 35-39.
O’Hanlon, R., A.R. McCracken, L.R. Cooke, 2016: Diversity and ecology of Phytophthora species on the island of Ireland. Biology and Environment: Proceedings of the Royal Irish Academy 116B (1), 27, DOI: 10.3318/bioe.2016.03.
Pirc, M., T. Dreo, D. Jurc, 2018: First Report of Pseudomonas syringae pv. aesculi as the Causal Agent of Bleeding Canker of Horse Chestnut in Slovenia. Plant Disease 102 (10), 2025, DOI: 10.1094/PDIS-12-17-1868-PDN.
Rosner, A., L. Maslenin, S. Spiegel, 1997: The use of short and long PCR products for improved detection of prunus necrotic ringspot virus in woody plants. Journal of virological methods 67 (2), 135-141.
Rust, S., 2013: Stress bei Bäumen. In: Roloff, A., 2013: Baumpflege. Stuttgart-Hohenheim, Eugen Ulmer Verlag. S. 68-76.
Schena, L., J.M. Duncan, D.E.L. Cooke, 2007: Development and application of a PCR-based ‘molecular tool box’ for the identification of Phytophthora species damaging forests and natural ecosystems. Plant Pathology 57, 64-75, DOI: 10.1111/j.1365-3059.2007.01689.x.
Schmidt, O., D. Dujesiefken, H. Stobbe, U. Moreth, R. Kehr, T. Schröder, 2008: Pseudomonas syringae pv. aesculi associated with horse chestnut bleeding canker in Germany. Forest Pathology 38 (2), 124–128, DOI: 10.1111/j.1439-0329.2007.00539.x.
Schmidt, O., U. Moreth, D. Dujesiefken, H. Stobbe, O. Gaiser, 2009: Fast molecular detection of Pseudomonas syringae pv. aesculi in diseased horse chestnut trees. Forest Pathology 39 (5), 343–348, DOI: 10.1111/j.1439-0329.2009.00595.x.
Schmidt, O., D. Dujesiefken, H. Stobbe, 2014: Infektionsversuche anRosskastanien mit Pseudomonas syringae pv. aesculi. Journal für Kulturpflanzen 66 (4), 130-135, DOI: 10.5073/JFK.2014.04.02.
Shigo, A.L., 1984: Compartmentalization: A Conceptual Framework for Understanding How Trees Grow and Defend Themselves. Annual Review of Phytopathology 22 (1), 189–214, DOI: 10.1146/annurev.py.22.090184.001201.
Werres, S., J., Richert, I., Veser, 1995: Untersuchungen von kranken und abgestorbenen Roßkastanien (Aesculus hippocastanum L.) im öffentlichen Grün. Nachrichtenbl. Deut. Pflanzenschutzd., 47 (4), 81-85.
Werres, S., S., Wagner, 2015: Absterbeerscheinungen an Rosskastanie – was sind die möglichen Ursachen?. Pro Baum 2015 (1), 18-22.
Werres, S., 2011: Phytophthora an Gehölzen – wie ist der aktuelle Stand? In: Dujesiefken, D. (Hrsg.), Jahrbuch der Baumpflege 2011: Yearbook of Arboriculture. Braunschweig, Haymarket Media. 159-168.

 Suchen
Suchen