
Fruchtfolgen mit und ohne Leguminosen: ein Review
Crop rotations with and without legumes: a review
Journal für Kulturpflanzen, 72 (10-11). S. 489–509, 2020, ISSN 1867-0911, DOI: 10.5073/JfK.2020.10-11.01, Verlag Eugen Ulmer KG, Stuttgart
 | Dies ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Namensnennung 4.0 International Lizenz (CC BY 4.0) zur Verfügung gestellt wird (https://creativecommons.org/licenses/by/4.0/deed.de). This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/deed.en). |
Leguminosen sind im ökologischen Landbau aufgrund ihrer Fähigkeit zur Luftstickstofffixierung unverzichtbar für die Stickstoffversorgung der Ackerkulturen und die Proteinversorgung der Nutztiere. Im konventionellen Anbau bieten Leguminosen die Möglichkeit, die häufig getreideintensiven Fruchtfolgen aufzulockern. Eine der wichtigsten Wirkungen dieses Break-crop-Effekts ist das Durchbrechen des Lebenszyklus von fruchtartenspezifischen Pathogenen und der damit verbundenen Einsparung von Pflanzenschutzmitteln. Die vorliegende Übersichtsarbeit stellt den derzeitigen Stand des Wissens zu Fruchtfolgen mit und ohne Leguminosen zusammen. Dabei werden ackerbauliche, ökologische und ökonomische Wirkungen des Anbaus groß- und kleinkörnig Leguminosenarten als Haupt- oder Zwischenfrüchte bzw. Untersaaten oder als Komponenten in Gemengen dargestellt und bewertet. Der Schwerpunkt liegt dabei auf relevanten Publikationen in wissenschaftlichen Journalen sowie Praxis- und Forschungsberichten der Jahre 2010 – 2020 die in Deutschland oder vergleichbaren klimatischen Bedingungen durchgeführt wurden. Abschließend wird daraus der notwendige Forschungsbedarf für die Themenbereiche Pflanzenbau (konventionell und ökologisch), Pflanzenschutz, Ökonomie, Ökologie und Klimaschutz abgeleitet.
Stichwörter: Leguminosen, Fruchtfolgen, Pflanzenbau, Pflanzenschutz, ökologischer Landbau, Biodiversität, Klimaschutz, Wirtschaftlichkeit, Forschungsbedarf
In organic farming, legumes are indispensable for the nitrogen supply of arable crops and the protein supply of livestock due to their ability to fix atmospheric nitrogen. In conventional farming, legumes offer the potential to break the often cereal-intensive crop rotations. One of the most important consequences of the break-crop effect is the interruption of the life cycle of crop-specific pathogens and the associated savings of pesticides. This review summarizes the current state of knowledge on crop rotations with and without legumes. It presents and evaluates the agronomic, ecological and economic effects of the cultivation of large- and small-seeded legume species as main or catch crops, when undersown in other crops, or used as components in mixtures. The focus is on relevant publications in scientific journals as well as practice and research reports published between 2010 and 2020 which were carried out in Germany or under comparable climatic conditions. Finally, essential research needs in the areas of crop production (conventional and organic), crop protection, economics, ecology, and climate protection are identified.
Key words: Legumes, crop rotations, plant production, plant protection, organic farming, biodiversity, climate protection, economics, research needs
In Anlehnung an Castellazzi et al. (2008) definieren wir Fruchtfolgen (Rotationen) als zyklisch wiederkehrende Abfolgen von Kulturpflanzenarten auf derselben Fläche. Fruchtfolgen sind zentraler Bestandteil von Anbausystemen. Letztere beinhalten darüber hinaus auch Managementaktivitäten wie beispielsweise Bodenbearbeitung, Aussaat, Düngung, Pflanzenschutz und Ernte (Vereijken, 1997). Über die Auswahl der Fruchtfolgefelder als kleinster Gliederungseinheit der Fruchtfolge entscheiden die Nachfrage der Verbraucher, Deckungsbeiträge, agrarpolitische Förder- und Marktordnungsmaßnahmen sowie zunehmend auch der Klimawandel. Seit längerer Zeit sind in Deutschland vereinfachte, weniger diverse Fruchtfolgen Realität, wobei der Anteil der Getreidearten, insbesondere der von Winterweizen (Triticum aestivum L.) und Mais (Zea mays L.), einseitig zu Lasten der Blattfrüchte zugenommen hat (Steinmann & Dobers, 2013; Stein & Steinmann, 2018). Die vier wichtigsten Kulturarten Winterweizen, Mais, Wintergerste (Hordeum vulgare L.) und Winterraps (Brassica napus L.) haben einen Anteil an der Ackerfläche von fast 70 % (Destatis, 2019a); in einigen Landkreisen kann der Anteil sogar deutlich darüber liegen. Die heute verbreiteten „simplifizierten“ Fruchtfolgen sind auch Ergebnis der Aufgabe des landwirtschaftlichen Gemischtbetriebs, d. h. der räumlichen und organisatorischen Trennung von Feld- und Viehwirtschaft (Köpke, 2013).
Es gibt viele pflanzenbauliche Gründe, diese engen Fruchtfolgen wieder mit Blattfruchtarten zu erweitern. Vielfältige Fruchtfolgen dienen u. a. dem Erhalt der Bodenfruchtbarkeit und der Nutzung phytosanitärer Effekte. So gehen mit zunehmender Getreidekonzentration Probleme einher wie das verstärkte Auftreten von Pilzkrankheiten und Ungräsern [z. B. Ackerfuchsschwanz (Alopecurus myosuroides Huds.) und Trespen (Bromus spp.)] sowie die zunehmende Resistenz von Schaderregern gegenüber Insektiziden und von Ungräsern gegenüber Graminiziden (Degner, 2013).
Für diese wünschenswerte Erweiterung der Fruchtfolgen bieten sich insbesondere Leguminosenarten an. Diese werden vorwiegend als Sommerungen angebaut. Es existieren auch Winterformen von Erbse, Ackerbohne und Lupine, die jedoch noch züchterisch besser an unsere Klimabedingungen angepasst werden müssen. Leguminosen werden vorwiegend in Form von Körnern oder Ganzpflanzen als Tierfutter eingesetzt. Bei den Körnerleguminosen verbleibt das Stroh auf dem Acker. Einige Arten besitzen ein tiefreichendes Wurzelsystem, verbessern die Bodenstruktur und gelten als humusmehrend (Rühl et al., 2009). Leguminosen sind aufgrund ihrer Fähigkeit zur Luftstickstofffixierung als Stickstoffinputquelle für den Betriebskreislauf sowie als Basis für die heimische Proteinversorgung in der Tierhaltung grundsätzlich unverzichtbar, insbesondere im ökologischen Landbau. Im konventionellen Anbau werden Leguminosen als eine Möglichkeit gesehen, die vorwiegend getreidebasierten Fruchtfolgen aufzulockern. Die gezielte Nutzung von Vorfruchteffekten bringt dem Landwirt vielfältige Vorteile (Christen, 2001; Andert et al., 2018; Henne et al., 2018).
Zahlreiche Untersuchungen weisen darauf hin, dass verschiedene ackerbauliche Probleme durch die Diversifizierung der Fruchtfolgen mit Leguminosen gelöst oder zumindest abgemildert werden könnten (DAFA, 2012). Für die Integration von Leguminosen in Fruchtfolgen werden als positive Aspekte neben der Bindung von Luftstickstoff und der Förderung der biologischen Vielfalt auch die Durchbrechung des Lebenszyklus von fruchtartenspezifischen Pathogenen, die Verfügbarmachung von Phosphor und anderer Nährstoffe sowie die Verbesserung der Bodenstruktur und des Wasser- und Lufthaushaltes des Bodens genannt (Cass et al., 2014; DAFA, 2012; Everwand et al., 2017; Finckh et al., 2015; Congreves et al., 2015; Steffens et al., 2005).
In neueren Studien konnten weitere Erkenntnisse zur phytopathologischen Situation einschließlich der Ableitung von Anbaupausen, Nährstoffversorgung und Düngung, Unkrautregulierung sowie zu neuen Anbauverfahren unter Berücksichtigung des Gemengeanbaus bei reduzierter Bodenbearbeitung bis hin zur Direktsaat nach entsprechenden Vorfrüchten gewonnen werden (Köpke et al., 2011; Wilbois et al., 2013; Böhm et al., 2014; Köpke et al., 2016). Die Ergebnisse aus weiterführenden Versuchen zum Gemengeanbau ermöglichten kulturart- und standortspezifische Empfehlungen für den ökologischen Landbau (Böhm et al., 2013). Allerdings weisen die Studien auch darauf hin, dass die meisten Leguminosen empfindlich auf Selbstfolgen reagieren („Leguminosenmüdigkeit“), weshalb größere Anbauabstände erforderlich sein können. Zudem treten einige Schaderreger sowohl bei Körner- als auch Futterleguminosen auf (Finckh et al., 2015).
Als Voraussetzung für die verstärkte Integration von Leguminosenarten in Fruchtfolgen müssen noch bestehende Wissenslücken geschlossen werden. Zum allgemeinen Forschungsbedarf bei Leguminosen existieren bereits verschiedene Stellungnahmen, welche nicht an Aktualität verloren haben. Dazu gehören das Fachgespräch „Anbau und Züchtung von Leguminosen in Deutschland“ (Wehling, 2009), das DAFA-Fachforum Leguminosen (DAFA, 2012) und der Bericht von Zerhusen-Blecher & Schäfer (2013). Studien, die auf den direkten Vergleich von Fruchtfolgen mit und ohne Leguminosen fokussieren und dabei einen oder mehrere Fruchtfolgezyklen umfassen, wurden bisher nicht veröffentlicht. Die Wirkungen des Anbaus von Leguminosen wurden zumeist nur für die direkte Folgekultur geprüft (Preissel et al., 2015; Cernay et al., 2018).
Ziel des vorliegenden Reviews ist es, einen Überblick über den derzeitigen Stand des Wissens zu Wirkungen der Integration von Leguminosen in Fruchtfolgen zu geben, die Befunde kritisch zu bewerten sowie bestehenden Forschungsbedarf abzuleiten. Dieser Review basiert auf einer vom Bundesministerium für Ernährung und Landwirtschaft (BMEL) im Jahr 2018 beim Thünen-Institut und Julius Kühn-Institut in Auftrag gegebenen Studie. In dieser Studie wurden ackerbauliche, ökologische und ökonomische Gesichtspunkte des Anbaus groß- und kleinkörnig Leguminosenarten als Haupt- oder Zwischenkultur bzw. Untersaat oder als Komponenten in Gemengen berücksichtigt. Der Schwerpunkt des Reviews liegt auf Studien und Forschungsansätzen, die in den letzten zehn Jahren durchgeführt wurden. Am Ende des Reviews wird der Forschungsbedarf für die Themenbereiche Pflanzenbau (konventionell und ökologisch), Pflanzenschutz, Biodiversität, Klimaschutz und Ökonomie aufgezeigt.
Trotz einiger Förderprogramme auf verschiedenen politischen Ebenen (Länder/Universitäten, Bund, EU) nahm die Anbaufläche von Körnerleguminosen in Deutschland und Europa in den vergangenen Jahren nach zwischenzeitlichen kurzfristigen Zunahmen immer wieder ab und liegt derzeit auf niedrigem Niveau. Seit Einführung des Greening hat sich der Anbau der Körnerleguminosen Ackerbohne (Vicia faba L.), Erbse (Pisum sativum L.), Lupine (Lupinus spp.) und Sojabohne [Glycine max (L.) MERR.] in Deutschland von 83.600 ha (2014) auf 174.000 ha (2019) mehr als verdoppelt (Destatis, 2015; Destatis, 2019b) (Abb. 1).
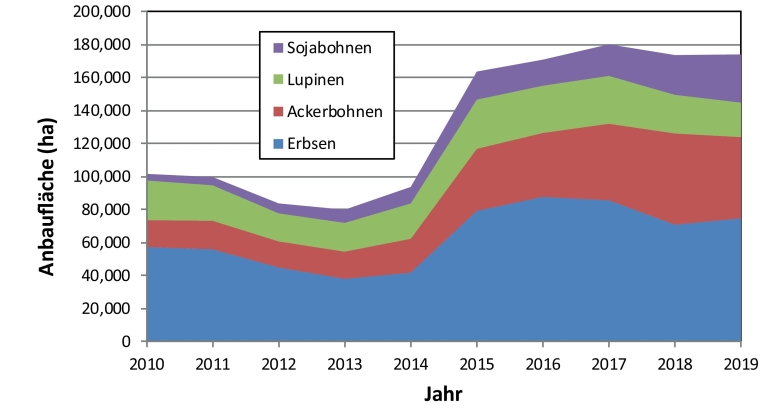
Abb. 1. Anbaufläche der Körnerleguminosen Futtererbse, Ackerbohne, Lupine und Sojabohne in Deutschland in den Jahren 2010 – 2019. Eigene Darstellung; Quelle: Statistisches Jahrbuch über Ernährung, Landwirtschaft und Forsten (verschiedene Jahrgänge), Hrsg.: Bundesministerium für Ernährung und Landwirtschaft.
Dennoch entspricht die gegenwärtige Leguminosenanbaufläche weniger als 1,5 % der gesamten Ackerfläche. Der Anteil an Körnerleguminosen im ökologischen Landbau nahm dagegen von 31 % im Jahr 2014 auf etwa 24 % im Jahr 2018 ab, wobei die Flächenentwicklung bei einzelnen Leguminosenarten sehr unterschiedlich verlief (Abb. 2). Diese Zahlen zeigen, dass der Flächenzuwachs insbesondere im konventionellen Bereich erfolgte.
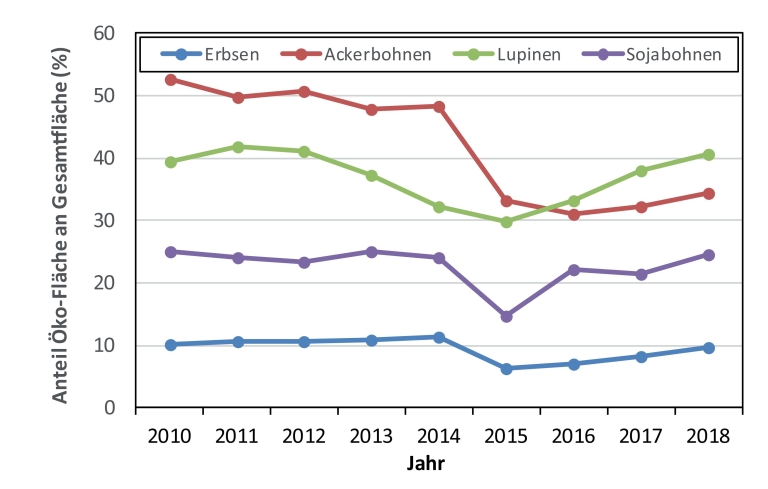
Abb. 2. Anteil ökologisch angebauter Körnerleguminosen an der jeweiligen Gesamtanbaufläche in Deutschland im Zeitraum von 2010 – 2018 . Eigene Darstellung; Quellen: Statistisches Jahrbuch über Ernährung, Landwirtschaft und Forsten (verschiedene Jahrgänge), Hrsg.: Bundesministerium für Ernährung und Landwirtschaft; AMI Markt Studie – Strukturdaten im ökologischen Landbau in Deutschland (verschiedene Jahrgänge), Hrsg.: Agrarmarkt Informations-Gesellschaft mbH (AMI); AMI Markt Bilanz Öko-Landbau (verschiedene Jahrgänge), Hrsg.: Agrarmarkt Informations-Gesellschaft mbH (AMI).
Nicht berücksichtigt wurden die im Gemenge angebauten Körnerleguminosen. Der Gemengeanbau in Deutschland umfasste im Jahr 2018 100.900 ha, wovon 41.000 ha auf den ökologischen Landbau entfielen (Schaack et al., 2019). Auch der Anbau von kleinkörnigen bzw. Futterleguminosen nahm in den letzten Jahren leicht von 246.000 ha im Jahr 2010 auf 283.300 ha im Jahr 2018 zu. Im Jahr 2018 standen davon 34 %, d. h. 96.000 ha, im ökologischen Landbau (Schaack et al., 2019).
Das zentrale Problem der Körnerleguminosen besteht darin, dass bei ihnen (mit Ausnahme der Sojabohne), im Gegensatz zu den etablierten Fruchtarten, die Lieferketten und Märkte nicht ausreichend entwickelt sind (Meynard et al., 2013). Aufgrund ihrer geringen Marktbedeutung werden Körnerleguminosen nicht oder nur unzureichend züchterisch bearbeitet, es mangelt an Innovationen und an einem im Hinblick auf die Integration in Fruchtfolgen notwendigen angepassten Stickstoffmanagement (Magrini et al., 2016). Ansätze gegenläufiger Trends sind allerdings auch zu erkennen. So war im Jahr 2017 in Deutschland ein neuer Höchststand der Anbaufläche für Körnerleguminosen zu verzeichnen. Die Anbauflächen von Ackerbohne, Sojabohne und Süßlupine (Abb. 3) nahmen leicht zu (Abb. 1). Klar erkennbar ist aber auch die Abhängigkeit des Leguminosenanbaus von agrarpolitischen Fördermaßnahmen. Nachdem aufgrund geänderter Förderbedingungen auf Greening-Flächen keine Pflanzenschutzmittel mehr eingesetzt werden durften, ging die Anbauflächen von Erbsen und Lupinen im Anbaujahr 2018 umgehend zurück.

Abb. 3. Sortenprüfung der Blauen Lupine.
Körnerleguminosen können im konventionellen Anbau problematisch werden, weil ihre N-Fixierleistung aufgrund der 2017 novellierten Düngeverordnung (DüV, 2017) und der Stoffstrombilanzverordnung (StoffBilV, 2017) als Stickstoffzufuhr in die Bilanz aufgenommen werden muss. Bei hohen Körnerleguminosenerträgen sind hohe Bilanzüberschüsse auszuweisen, wodurch sich die gesamte Fruchtfolgebilanz verschlechtert (Pahlmann et al., 2018).
Eine Sonderstellung unter den Körnerleguminosen nimmt die Sojabohne ein. Obgleich ihrem Anbau in Europa klimatische Grenzen gesetzt sind, wird der Sojabohne, insbesondere nicht gentechnisch veränderten Sorten, ein beträchtliches Flächenentwicklungspotenzial zugeschrieben (De Visser et al., 2014). So wurden und werden neue Sorten entwickelt, die auch in kühleren Regionen mit kürzerer Vegetationszeit und geringeren Wärmesummen angebaut werden können (Zimmer et al., 2016).
Im ökologischen Landbau sind Leguminosen aufgrund ihrer Fähigkeit zur Luftstickstofffixierung als Stickstoffinputquellen für den Betriebskreislauf unverzichtbar. Insbesondere Futter- und Körnerleguminosen bilden die Basis für die Proteinversorgung in der Tierhaltung. Hieraus resultieren in der Regel vielseitige und mehrfeldrige Fruchtfolgen mit Leguminosen im ökologischen Landbau. Deren Bedeutung in der Fütterung führt außerdem zu einer höheren ökonomischen Bewertung der Leguminosen im Vergleich zum konventionellen Anbau. Hohe Anteile an Leguminosen in Fruchtfolgen können aber auch zu phytopathologischen Problemen führen. Im ökologischen Landbau geht es somit nicht um die Frage „Fruchtfolgen mit oder ohne Leguminosen“, sondern um die Wirkung von Leguminosen auf nicht-legume Folgefrüchte sowie eine nachhaltige Integration von Leguminosen in Fruchtfolgen ohne negative phytopathologische Nebenwirkungen.
In den Jahren 2010 – 2020 wurden vielfältige pflanzenbauliche Studien mit unterschiedlichen Leguminosenarten durchgeführt und neu begonnen. Neben spezifischen Untersuchungen an ausgewählten Arten in Reinsaat und Gemenge sind insbesondere die Demonstrationsnetzwerke Sojabohne (www.sojafoerderring.de), Erbse/Bohne (www.demoneterbo.agrarpraxisforschung.de) und Lupine (www.lupinen-netzwerk.de) hervorzuheben, welche u. a. den Wissens- und Erfahrungsaustausch zwischen Forschung, Beratung und Praxis fördern sollen. An diese Demonstrationsnetzwerke wurden Forschungs- und Entwicklungsvorhaben wie z. B. das BLE-geförderte Projekt RELEVANT (Regulierende Ökosystemleistungen in Fruchtfolgen mit Ackerbohne und Erbse: Quantifizierung, Bewertung und Realisierung; vgl. Kapitel Biodiversität) angegliedert (Schulz et al., 2019).
Im Rahmen interdisziplinär ausgerichteter Verbundvorhaben wurden wichtige Fragestellungen des Körnerleguminosenanbaus im ökologischen Landbau aufgegriffen und bearbeitet (Köpke et al., 2011; Wilbois et al., 2013; Böhm et al., 2014; Köpke et al., 2016). So konnten temporäre Direktsaatverfahren für Ackerbohnen in Hafer-Strohmulch etabliert werden, wobei diese bei geringem Druck durch perennierende Unkrautarten ohne wirtschaftliche Ertragseinbußen möglich sind (Köpke et al., 2011). Auch Untersuchungen zur reduzierten Bodenbearbeitung in Gemengeanbausystemen mit Wintererbsen (Abb. 4) zeigten keine Unterschiede in der Unkrautunterdrückung im Vergleich zur Pflugbearbeitung. Die untersuchten Gemengevarianten wiesen im Vergleich zu den Reinsaaten mit Wintererbsen einen deutlich geringeren Blattlausbefall auf (Gronle et al., 2014). Dies konnte auf geringe N-Gehalt der im Gemenge angebauten Erbsen zurückgeführt werden. Dabei lieferte der Gemengeanbau einer normalblättrigen Sorte höhere Erträge als eine halbblattlose Sorte in Reinsaat und im Gemenge (Gronle et al., 2015).

Abb. 4. Gemengeanbau von Weißer Wintererbse und Wintertriticale.
Untersuchungen an Körner- und Futterleguminosen zeigten, dass eine Schwefeldüngung bei Körnerleguminosen aufgrund des relativ geringen Schwefelbedarfes nur selten einen positiven Effekt auf Ertrag und Qualität hat (Schmidtke & Lux, 2015; Gruber & Wegner, 2017). Futterleguminosen reagieren deutlich positiv auf die Schwefeldüngung, weil die N-Fixierleistung erhöht und damit der Vorfruchtwert gesteigert wird (Fischinger et al., 2011; Becker et al., 2013; Böhm, 2016).
Für die pflanzenbauliche Beurteilung der Wirkung einer Eingliederung von Leguminosen in Fruchtfolgen ist es zweckmäßig, zwischen dem Stickstoffeffekt und dem Break-crop-Effekt zu unterscheiden (Jensen, 2006). Beide Effekte zusammen machen den Vorfruchtwert von Körnerleguminosen aus. Im Gegensatz zum leguminosenspezifischen Stickstoffeffekt ist der Break-crop-Effekt nicht an Leguminosen gebunden, sondern Ergebnis der Unterbrechung monotoner Fruchtfolgen. Man könnte den Break-crop-Effekt deshalb auch als Diversifizierungseffekt bezeichnen, welcher sich auch mit Blattfrüchten wie z. B. Raps erzielen lässt. Eine der wichtigsten Folgen des Break-crop-Effekts ist das Durchbrechen des Lebenszyklus von fruchtartenspezifischen Pathogenen, wodurch eine Einsparung von Pflanzenschutzmitteln ermöglicht wird (Munier-Jolain & Collard, 2006). Besonders groß sind die Vorteile der Einführung von Körnerleguminosen in Anbausystemen mit einem hohen Getreideanteil. Dagegen lässt sich in Regionen mit bereits vielgestaltigen Fruchtfolgen (z. B. in der Schweiz) durch die Aufnahme von Körnerleguminosen in die Fruchtfolge häufig überhaupt kein Break-crop-Effekt erzielen (Nemecek et al., 2008). Es gibt auch Hinweise darauf, dass die direkte Vorfrucht einen größeren Einfluss auf die Ertragshöhe der Nachfrucht hat als die gesamte vorangegangene Rotation, dass also der Vorfruchteffekt größer ist als die Fruchtfolgewirkung (Greef et al., 2004).
Die positive Wirkung von Körnerleguminosen auf die Erträge der Nachfrüchte bestätigen zwei neuere Meta-Studien (Preissel et al., 2015; Cernay et al., 2018). Die Auswertung des insgesamt 15 Länder umfassenden weltweiten Datenmaterials aus internationalen Peer-review-Journalen ergab, dass der Ertrag von Getreide nach Körnerleguminosen durchschnittlich um 29 % höher war als der Ertrag von Getreide nach Getreide (Cernay et al., 2018). Die positive Wirkung von Körnerleguminosen nahm allerdings mit zunehmender Stickstoffdüngung zur Getreidenachfrucht ab und war oberhalb von 150 kg N ha–1 sogar vernachlässigbar gering. Dieser kritische Wert von 150 kg N ha–1 wird in konventionellen europäischen Getreideanbausystemen häufig überschritten. Auch Preissel et al. (2015) zeigten erhöhte Erträge von Getreide nach Körnerleguminosen von 0,5 bis 1,6 t ha–1, wobei die Mehrerträge durch eine Leguminosenvorfrucht ebenfalls bei niedriger Stickstoffdüngung zu den nachfolgenden Kulturen am höchsten ausfielen. Dieser Mehrertrag war jedoch nur geringfügig größer als bei Wahl einer anderen Blattvorfrucht wie Raps oder Sonnenblume. Prinzipiell ließen sich also durch die Einführung von Körnerleguminosen in Fruchtfolgen hohe Getreideerträge auch bei deutlich reduziertem Stickstoffeinsatz erzielen.
Braun et al. (2014) verglichen unterschiedlich umfangreiche Fruchtfolgen mit und ohne Leguminosen und Zwischenfrüchten (einen Marktfruchtbetrieb, einen Veredlungsbetrieb mit Biogasanlage sowie einen optimierten Klimabetrieb mit Biogasanlage) hinsichtlich ihrer Wirkung auf das Klima. Bei der erweiterten Fruchtfolge des optimierten Klimabetriebs ergaben sich für die dort angebaute Wintergerste deutliche Ertragsvorteile durch die Vorfrucht Ackerbohne (Röper et al., 2017). Hier ist allerdings zu beachten, dass die Ackerbohne erhöhte Herbst-Nmin-Werte hinterlässt. In einem Langzeitversuch (1988 – 2001) der Christian-Albrechts-Universität zu Kiel wurden in Norddeutschland insgesamt 15 verschiedene Fruchtfolgen verglichen, welche Winterraps, Winterweizen, Wintergerste, Erbsen und Hafer (Avena sativa L.) beinhalteten (Sieling & Christen, 2015). Im Anschluss an den Fruchtfolgevergleich wurde zweimal Winterweizen angebaut, um die Wirkung der Vorfrüchte bzw. der Fruchtfolgen auf Wachstum, Ertrag und Ertragskomponenten zu quantifizieren. Die höchsten Weizenerträge wurden im ersten Jahr nach Erbse und Winterraps erzielt. Im zweiten Jahr unterschieden sich die Kornerträge aller Varianten nicht mehr signifikant voneinander.
Schneider et al. (2012) führten im Bereich des ökologischen Landbaus Fruchtfolgeversuche durch mit dem Ziel, Fruchtfolgen mit unterschiedlichen Leguminosenarten und deren Nutzung im Hinblick auf die Ertragsleistungen (inklusive Fruchtfolgeertrag und ökonomischer Bewertung) und die Qualitäten der nach Leguminosen angebauten nicht-legumen Fruchtarten zu vergleichen. Dabei wurde festgestellt, dass auf Kleegras basierende Fruchtfolgen mit Schnittnutzung (viehhaltende Systeme) die höchste Produktivität auf Basis von Getreideeinheiten erreichen. Beim Vergleich von Fruchtfolgen mit Körnerleguminosen ohne innerbetriebliche Verwertung und Fruchtfolgen mit Kleegrasanbau (gemulcht mit Verbleib auf der Fläche) zeigten sich letztere im Hinblick auf den Ertrag der nachgebauten Marktfrüchte überlegen (Schneider et al., 2012). Die Gesamtproduktivität der beiden Systeme wies indessen keine Unterschiede auf.
Im Rahmen eines 50 Jahre währenden Fruchtfolgeversuchs in Kanada wurden die N2O-Emissionen und der Maiskornertrag über drei Vegetationsperioden gemessen (Drury et al., 2014). In Rotation mit Luzerne war der Maiskornertrag im Durchschnitt fast doppelt so hoch wie der in der Mais-Selbstfolge; in beiden Varianten wurden gleiche N-, P- und K-Mengen als Startdünger ausgebracht und in die oberen 10 cm Bodenschicht eingearbeitet. Anhand von Ertrags- und Wetterdaten (1982 – 2012) aus einem weiteren kanadischen Dauerversuch untersuchten Gaudin et al. (2015) die Ertragsstabilität einer Mais-Monokultur sowie von Fruchtfolgen mit Sojabohne und Luzerne als Hauptfrucht und Getreide mit Kleeuntersaaten. Obwohl die Größenordnung der Fruchtfolgevorteile je nach Kultur, Wetterbedingungen und praktizierter Bodenbearbeitung variierte, nahm die Ertragsstabilität von Mais und Sojabohnen in den vielfältigeren Fruchtfolgen signifikant zu. Die Vorteile der diversifizierten Fruchtfolge kamen besonders unter ungünstigen Wetterbedingungen (kühl-feucht, trocken-heiß) zum Tragen. Insgesamt zeigte die Studie, dass vielfältigere Fruchtfolgen zu einer erhöhten Ertragsstabilität von Getreide beitragen.
Ein weiteres langjähriges und dreiortiges Fruchtfolgeexperiment wurde 1997 in Dänemark mit zwei ökologischen und einer konventionellen Fruchtfolge mit und ohne Einsatz von anaerob behandelter Gülle, Gründüngung (Luzerne oder Grasmischung mit Rot- und Weißklee) und Zwischenfrüchten (u. a. Erbse) begonnen (De Notaris et al., 2018). Für die ersten drei Fruchtfolgezyklen (1997 – 2008) zeigte die Fruchtfolge Sommergerste – Kleegras (gemulcht) – Kartoffel – Weizen im dritten Zyklus mit integrierter Gründüngung an allen Standorten tendenziell zunehmende Trockenmasseerträge (Olesen et al., 2011; Shah et al., 2017). Der Minderertrag des ökologischen gegenüber dem konventionellen Anbausystem betrug im dritten Zyklus zwischen 21 % und 64 %. Durch den Einsatz von Wirtschaftsdüngern ließen sich diese Unterschiede am stärksten reduzieren. Zusätzlich konnten eine ganzjährige Gründüngung (Kleegras, gemulcht) und leguminosenbasierte Zwischenfruchtgemenge die Ertragsleistungen verbessern. Mit dem Anbau einer ganzjährigen Gründüngung geht jedoch der Verlust einer Marktfrucht einher, sodass die Gesamtproduktivität des Anbausystems nicht erhöht wurde.
Während die Wirkung von Leguminosen auf den Ertrag der Folgekultur relativ einfach gemessen werden kann, sind andere Effekte, wie z. B. der Schädlings- und Pathogendruck, Veränderungen der Bodenstruktur oder des Wurzelwachstums nur schwer zu quantifizieren. In einem Langzeitversuch wurde eine geringere Humusreproduktionsleistung von Körnerleguminosen im Vergleich zu Kleegras bestätigt, während hinsichtlich Humusgehalt und Humusqualität zwischen der Form der Kleegrasnutzung (Schnitt mit Abfuhr und Rückführung über organische Düngung vs. Mulchen) keine Unterschiede auftraten (Urbatzka & Beck, 2015).
Congreves et al. (2015) werteten vier Langzeitfruchtfolgen mit Luzerne, Sojabohne und Klee als Untersaaten in Ontario (Kanada) hinsichtlich der Effekte auf die Bodengesundheit aus. Die Aggregatstabilität und der Gehalt an organischer Substanz wurden durch Luzerne gefördert. Demnach scheint der Einfluss einzelner Kulturen innerhalb einer Fruchtfolge für die Bodengesundheit entscheidender zu sein als eine möglichst große Diversität (Congreves et al., 2015).
Hinzu kommen die Effekte, die auf der Verfügbarmachung von anderen Nährstoffen als Stickstoff basieren. So können Hülsenfruchtarten wie Weiße Lupine, Kichererbse oder Ackerbohne über die Wurzelausscheidung von organischen Säuren schwer verfügbaren Phosphor mobilisieren (Hocking, 2001, Steffens et al., 2005) und die Versorgung der Folgekulturen mit pflanzenverfügbarem Phosphor verbessern (Nuruzzaman et al., 2005). Leguminosenarten mit Pfahlwurzeln können zudem die Bodenstruktur verbessern sowie den Boden mit organischem Kohlenstoff anreichern (Rochester et al., 2001; Aslam et al., 2003) und damit der Folgekultur einen größeren Wurzeltiefgang ermöglichen (Kahnt, 2008).
Das EU-Projekt Legume Root Impact beschäftigte sich mit dem zeitlichen Verlauf der Zersetzung von Wurzelsystemen von Futtermischungen (Rot- und Weißklee) im Boden. Dabei wurde festgestellt, dass die schneller abbaubare Blattmasse eine deutlich größere Rolle für den kurzfristigen Kohlenstoff- und Stickstoffumsatz im Boden spielt als die Wurzelmasse (CORDIS, 2014).
Neben diesen direkt auf Fruchtfolgewirkungen ausgerichteten Untersuchungen existieren Projektergebnisse aus Arbeiten zur optimalen Einbindung klein- und großkörniger Leguminosen in Fruchtfolgen. Diese enthalten Aussagen zur Vor- und Zwischenfruchtstellung auf verschiedenen Bodenarten und Angaben zu Möglichkeiten der organischen Düngung zu unterschiedlichen Leguminosenarten (Kolbe, 2009; Schlathölter, 2015; Schlathölter & Petersen, 2015; Böttcher & Schmidt, 2016; Pahlmann & Kage, 2018; Stute & Schäfer, 2018). Mit dem Anbau legumer Winterzwischenfrüchte oder Zwischenfruchtmischungen können Nährstoffe vor Auswaschung geschützt und Unkräuter unterdrückt werden. Von Vorteil ist dabei die Verwendung abfrierender Zwischenfrüchte. Soll der Frühjahrsaufwuchs genutzt werden, sind mögliche Einschränkungen in der Wasserversorgung für die Zweitfrucht zu berücksichtigen (Böttcher & Schmidt, 2016). Mehrfeldrige Fruchtfolgen können positive Effekte auf Stickstoffeffizienz und -bilanz ausüben, erschweren jedoch eine bedarfsgerechte Stickstoffdüngung für einzelne Fruchtfolgefelder.
Als ein Nachteil von Fruchtfolgen mit Körnerleguminosen führen Nemecek et al. (2008) das erhöhte Nitratauswaschungspotential an, welches u. a. mit der symbiontischen Stickstofffixierung bzw. der schwer kalkulierbaren Mineralisierung von organisch gebundenem Stickstoff zusammenhängt. Nach Ansicht der Autoren lässt sich dieses Problem durch den Einsatz von Zwischenfrüchten, Winter-Körnerleguminosen oder einen Gemengeanbau reduzieren.
Auch eine mehrjährige dänische Studie mit zwei ökologischen und einer konventionellen Fruchtfolge mit und ohne Einsatz von anaerob behandelter Gülle, Gründüngung [Luzerne oder Grasmischung mit Rotklee (Trifolium pratense L.) und Weißklee (Trifolium repens L.)] und Zwischenfrüchten (u. a. Erbse) weist Ergebnisse zur Nitratauswaschung aus (De Notaris et al., 2018). Sowohl in den konventionellen als auch in den ökologischen Systemen konnten legume und nicht-legume Zwischenfrüchte die Stickstoffauswaschung im vierten Zyklus des Fruchtfolgeexperiments um durchschnittlich 60 % reduzieren. Beim ökologischen Anbau mit Körnerleguminosen erhöhte der Einsatz von Zwischenfrüchten auch die Stickstoffproduktion um jährlich durchschnittlich 20 kg N ha–1. Das Stickstoffauswaschungsrisiko war beim ökologischen Anbau mit Gründüngung (2-jährig gemulchtes Kleegras) höher als bei den anderen Anbausystemen.
Neben den unterschiedlichen Wirkungen von Leguminosen auf Folgekulturen ließen sich umgekehrt auch Wirkungen von Nicht-Leguminosen auf Leguminosen in Gemenge oder als Vorfrucht nachweisen (CORDIS, 2015). Im Rahmen des EU-Projekts PROLEGSO konnten die Leguminosen Rotklee, Weißklee und Hornklee (Lotus spp.) in Form von höheren Erträgen von Nicht-Leguminosen wie Deutschem Weidelgras (Lolium perenne L.), Ruchgras (Anthoxanthum odoratum L.), Spitzwegerich (Plantago lanceolata L.) und Gewöhnlicher Schafgarbe (Achillea millefolium L.) profitieren, die langsam wachsen und wenig Phosphor und Kalium aufnehmen. Derartige Nicht-Leguminosen könnten den Boden für Leguminosen durch Förderung symbiontischer Organismen (Mycorrhiza-Pilze, Rhizobien) verbessern.
Der Integration von Leguminosen in Fruchtfolgen sind allerdings Grenzen gesetzt, da einige Arten aufgrund einer Selbstunverträglichkeit empfindlich auf einen zu häufigen Anbau reagieren. So konnten Schmidt et al. (2014) in Untersuchungen auf ökologisch bewirtschafteten Praxisbetrieben zeigen, dass der Kornertrag von weißblühenden Erbsen mit zunehmender Anbauhäufigkeit abnimmt. Sie leiteten aus den Ergebnissen für weißblühende Erbsen einen Anbauabstand von mindestens 9 – 10 statt der bislang empfohlenen 5 – 6 Jahre ab. Grund dafür sind die zum sogenannten Ascochyta-Komplex der Erbse (Fuß- und Brennfleckenkrankheiten) gehörenden bodenbürtigen Schaderreger wie z. B. Mycoshaerella pinoides und Phoma medicaginis (Kraft & Pfleger, 2001; Finckh et al., 2015), die im Boden zum Teil eine Überlebensdauer von mehr als zehn Jahren haben. Nach Bretag et al. (2001) können sie Ertragsverluste von bis zu 75 % verursachen. Zudem können ein Teil dieser und anderer Schaderreger sowohl an Körner- als auch an Futterleguminosen auftreten (Finckh et al., 2015). Die derzeit empfohlenen Anbauabstände für die am häufigsten angebauten Leguminosenarten zeigt Tab. 1.
Tab. 1. Empfohlene Anbaupausen für verschiedene Leguminosenarten. Quellen: DemoNetErbseBohne (2020), KTBL (2013).
Futterleguminosen |
| Körnerleguminosen | ||
Art | Jahre |
| Art | Jahre |
Seradella | 1 – 2 |
| Sojabohne | 1 – 3 |
Weißklee | 1 – 3 |
| Ackerbohne, Lupine, Wicke, Linse | 5 – 7 |
Alexandrinerklee, Perserklee | 3 – 4 |
| Grünfuttererbse (Peluschke) | 5 – 9 |
Rotklee, Schwedenklee, Luzerne, Esparsette, Gelbklee, Inkarnatklee | 4 – 7 |
| Erbse | 6 – 10 |
Weitere Erkenntnisse zu Fruchtfolgewirkungen von Leguminosen sind zukünftig aus den im Rahmen des Programms Horizon 2020 gestarteten neuen EU-Projekten zu erwarten. Diese beschäftigen sich mit der Schaffung der Voraussetzungen für erfolgreiche Anbausysteme unter Nutzung von Klee (Trifolium spp.), Ackerbohne, Linse (Lens culinaris MEDIK.), Lupine, Sojabohne, Kichererbse (Cicer arietinum L.), Bohne (Phaseolus spp.), Körnererbse, Luzerne (Medicago sativa L.) und Augenbohne [Vigna unguiculata (L.) WALP.]. Einen weiteren Schwerpunkt bildet die Ermittlung der Vorteile von Artenmischungen unter Einbeziehung von Leguminosen wie Körnererbse, Klee, Bockshornklee (Trigonella foenum-graecum L.) und Wicke (Vicia spp.) zur Entwicklung diversifizierter und belastbarer Anbausysteme, die weniger von externen Inputs abhängig sind (EU-Projekt ReMIX). Vielversprechend ist auch das im November 2018 gestartete Thematic Network (H2020) Legumes Translated. Dieses hat zum Ziel, bestehendes Wissen zum Anbau von Körnerleguminosen zu sammeln, Innovatoren im landwirtschaftlichen Bereich mit Wissenschaftlern zusammenzubringen und gemeinsam Praxisempfehlungen für den Anbau von Körnerleguminosen zu entwickeln (Murphy-Bokern et al., 2019; CORDIS, 2020).
Die erfolgreiche Produktion von Leguminosen hängt von der Verfügbarkeit resistenter Sorten und der hinreichenden Verfügbarkeit von Pflanzenschutzmitteln und -verfahren, der Überwachung der Zielorganismen sowie der Feldhygiene ab (Stoddard et al., 2010). Während für großflächig angebaute Kulturen hocheffiziente Pflanzenschutzmittel zur Verfügung stehen, ist die entsprechende Verfügbarkeit für Leguminosen eher eingeschränkt. Vergleichende Studien zu mitteleuropäischen Fruchtfolgen mit und ohne Leguminosen im Hinblick auf das Auftreten von Schädlingen sind kaum verfügbar. Anhaltspunkte hierfür können häufig nur aus internationalen Studien abgeleitet werden (Sillero et al., 2010). Das Auftreten von typischen Leguminosenschädlingen wird durch die Häufigkeit des Leguminosenanbaus in einer Region beeinflusst, aber auch durch natürlich in Grünlandflächen und an Ackerrändern vorkommende Leguminosen. Andere ackerbauliche Kulturen als Leguminosen werden von speziellen Leguminosenschädlingen im Allgemeinen nicht befallen.
Der Review von Sillero et al. (2010) zum Auftreten von Schadpathogenen sowie den Möglichkeiten der Bekämpfung kommt zum Ergebnis, dass beim Anbau von großkörnigen Leguminosen häufig die praktizierte Fruchtfolge und die Jahreswitterung in Verbindung mit den eingesetzten Bodenbearbeitungsverfahren maßgeblich für das Befallsauftreten verantwortlich sind. In Fruchtfolgen mit Zuckerrüben (Beta vulgaris ssp. vulgaris) oder Raps besteht die Gefahr, dass bodenbürtige Schaderreger wie Rhizoctonia spp. und Sclerotinia spp. im Bodensubstrat angereichert werden (Noack, 2016). Aufgrund ihres weiten Wirtsspektrums können diese Schadorganismen zu einem kulturartübergreifenden Problem für die gesamte Fruchtfolge werden. Daher gilt es, entsprechende Anbaupausen einzuhalten (vgl. Tab. 1).
Die Verbreitung oder Übertragung von Fruchtfolgekrankheiten werden bei der Sojabohne derzeit als weniger bedeutsam eingeschätzt. Mit Diaporthe spp. und Phomopsis spp. sind zwei wichtige Schadpilze aus anderen Anbauregionen der Welt bekannt (Xue et al., 2007), die bei der Sojabohne im Falle eines größerflächigen Anbaus erhebliche Schäden an Stängel, Hülsen und Samen verursachen können. Der Aufgang und die Etablierung von Sojabohnenbeständen lassen sich derzeit durch den Einsatz von Saatgutbehandlungen mit den Wirkstoffen Fludioxonil, Metalaxyl, Difenoconazol und Trifloxystrobin sowie dem biologischen Präparat Clonostachys rosea verbessern (Xue et al., 2007). Einige dieser Wirkstoffe werden aber aufgrund einer Neubewertung durch die EU-Kommission nach der Verordnung 1107/2009/EU in Zukunft nicht mehr verfügbar sein. Nach Angaben von Shuxian & Chen (2012) sind Resistenzquellen für die Züchtung verfügbar.
Versuche zur Bekämpfung der o. g. Krankheiten mit biologischen Mitteln, Resistenzinduktoren und Pflanzenextrakten hatten einen lediglich unbefriedigenden oder gar keinen Erfolg (Stoddard et al., 2010). Demgegenüber gibt es erfolgversprechende Ansätze, dass Anthraknose (Abb. 5), hervorgerufen durch Colletotrichum spp., biologisch durch Clonostachys spp., Bacillus subtilis oder Pseudomonas putida kontrolliert werden kann (Tinivella et al., 2009).

Abb. 5. Anthraknose an Weißer Lupine.
Nach Studien von Hein (2006) ist der Verlust durch tierische Organsimen höher zu bewerten als der durch Schadpilze. Neben Schäden durch einen direkten Befall der Kulturpflanze muss auch die Funktion der Schädlinge als Virusvektoren (z. B. PNYDV übertragen durch Acyrthosiphon pisum) berücksichtigt werden (Männel et al., 2018).
Ursachen für zunehmende wirtschaftliche Schäden ist die fehlende Verfügbarkeit sowohl von wirksamen Insektiziden als auch von toleranten oder resistenten Sorten. Nach derzeitigem Kenntnisstand wird die Zunahme der Insektizidresistenz (z. B. bei Blattläusen gegenüber Pyrethroiden) die Möglichkeiten einer gezielten Bekämpfung weiter deutlich mindern.
Ein spezifischer Schaderreger der Erbse ist der Erbsenwickler Cydia nigricana. Ein höherer Anteil an Erbsenfeldern in der Landschaft fördert das Auftreten dieses Schaderregers (Huusela-Veistola & Jauhiainen, 2006). Dies gilt auch für weitere Leguminosenschädlinge, wenn der Anteil der betroffenen Leguminosen in der Fruchtfolge zunimmt. Dabei ist zu berücksichtigen, dass die Schwarze Bohnenblattlaus (Aphis fabae) neben der Ackerbohne auch Nichtleguminosen wie die Rübe und verschiedene Wildpflanzen befällt, jedoch keine Erbsen.
Kleinkörnige Leguminosen werden ebenfalls von spezifischen Schadinsekten wie verschiedenen Samenkäfern (Bruchidae), Blattrandkäfern der Gattung Sitona und Blattnagern der Rüsselkäfergattung Hypera befallen. Polyphage Blattlausarten wie Myzus persicae können als Virusüberträger in verschiedenen Kulturen relevant sein. Bei länger auf der Fläche stehenden kleinkörnigen Leguminosen (Klee, Luzerne) können Drahtwürmer (Larven der Elateridae) gefördert werden.
Geprüft wurden in einigen Untersuchungen auch die Auswirkungen des „intercropping“, also dem Gemengeanbau von Körnerleguminosen und z. B. Getreidearten. Dabei konnte vielfach eine Reduzierung des Blattlausbefalls (Myzus persicae) in den Gemengevarianten (Seidenglanz et al., 2011; Bedoussac, 2009; Gronle et al., 2014), bei Erbsenwicklern (Cydia nigricana) hingegen keine Unterschiede (Gronle et al., 2014) festgestellt werden.
Die Auswertung verfügbarer Studien in der Bekämpfung von Unkräutern und Ungräsern zeigt, dass dieser Bereich bei der Etablierung der Leguminosen in einer betriebsüblichen Fruchtfolge einen wesentlichen Einfluss hat. Insbesondere bei der Bekämpfung von Ungräsern haben die Körnerleguminosen aufgrund ihres langsamen Wachstums nur ein geringes Unkrautunterdrückungsvermögen (Böhm, 2014). Die Verfügbarkeit geeigneter Mittel ist sehr eingeschränkt und wird in manchen Praxisbetrieben als Ausschlusskriterium für den Anbau von Leguminosen gesehen. Studien der Offizialberatung zeigen, dass mechanische Verfahren wie Blindstriegeln, Blindeggen und Reihenhacken gerade unter trockenen Bodenbedingungen sehr effektiv sind (Abb. 6). Ein Teil der Maßnahmen kann unter Beachtung der Saatgutablagetiefe bereits vor dem Feldaufgang erfolgen und so Wirkungslücken nutzbarer Herbizide schließen. Hackgeräte in Kombination mit optischen Verfahren und digitalen Erfassungs- und Steuerungseinheiten können höhere Wirkungsgrade erzielen (z. B. Gerhards et al., 1998; Heinold et al., 2018; Heuser et al., 2018).

Abb. 6. Möglichkeiten der mechanischen Unkrautbekämpfung in Leguminosen. Links: Einsatz eines Rollstriegels in Blauen Lupinen; rechts: Einsatz eines Hackstriegels in Futtererbsen.
Die Nutzung von Körnerleguminosen wie Sojabohne oder Ackerbohne und Futterleguminosen wie Luzerne und diverse Kleearten in ackerbaulichen Fruchtfolgen hat potenziell vielfältige Auswirkungen auf Ackerwildkräuter, Insekten und Wirbeltiere und damit auf die Biodiversität in Agrarlandschaften (Cass et al., 2014; Everwand et al., 2017). Generell bedeutet jede zusätzliche die Fruchtfolge erweiternde Ackerfrucht eine Erhöhung des Artenspektrums und damit der Komponente der genetischen Vielfalt innerhalb der Agrobiodiversität. Des Weiteren haben Leguminosen spezifische Eigenschaften, welche sowohl ober- als auch unterirdisch Komponenten der Biodiversität in ihrer Umgebung fördern können. Dazu gehören die Fähigkeit zur biologischen N-Fixierung, ein enges C/N-Verhältnis der Pflanzenbiomasse (Aher et al., 2017), extraflorale Nektarien (Free, 1962) und eine von Getreide abweichende Bestandsarchitektur (Abbo et al., 2009). Durch ein großes Blütenangebot können Leguminosen, welche zur Blüte gelangen, ebenso wie Raps oder andere Massentrachten auch, generalistische Bestäuberarten in Agrarlandschaften fördern (Westphal et al., 2003). Dabei sind Leguminosen in ihren Nutzungsmöglichkeiten aber häufig vielseitiger als andere Kulturen. Neben dem Einsatz als Hauptfrucht können Leguminosen auch als Gründüngung, Zwischenfrucht und als Partner im Mischanbau vor allem mit Getreidekulturen (Erbse/Gerste, Ackerbohne/Weizen u. a.) fungieren. Dadurch können sie sowohl zu einer räumlichen als auch zeitlichen Diversifizierung der Fruchtfolgen beitragen, woraus differenzierte Einflüsse auf die Biodiversität resultieren können. Im Folgenden gehen wir auf Auswirkungen des Leguminosenanbaus auf bestäubende Insekten, räuberische und pflanzenfressende Arthropoden sowie Regenwürmer ein.
Trotz der bekannten Tragweite der Bestäubung als Ökosystemleistung (Gallai et al., 2009), gibt es immer mehr Beweise für den Rückgang der Bestäuberzahlen auf der ganzen Welt (Potts et al., 2010). Dieser Rückgang steht u. a. im Zusammenhang mit dem Verlust von Lebensräumen und Nahrungsressourcen (Goulson et al., 2015). Blühende Leguminosen (Abb. 7), welche eine Nahrungsgrundlage für nektarsammelnde und bestäubende Insekten in Agrarlandschaften bieten, können hier förderlich wirken (Woodcock et al., 2014). Futterleguminosen wie zum Beispiel Klee, Esparsette (Onobrychis spp.), Wicken und Luzerne stellen Bestäubern ein reichhaltiges Angebot an Blüten zur Verfügung. Das Blühangebot von Körnerleguminosen hingegen ist geringer, sie lassen sich aber als Trachtpflanzen gezielt in Fruchtfolgen einbauen (Ministerium für Ländlichen Raum und Verbraucherschutz, 2019). Trachtpflanzen sind solche Pflanzen, die besonders reichhaltig an Nektar und Pollen sind und deshalb von Bienen für die Erzeugung von Honig bevorzugt werden. Um Trachtlücken zu schließen, eignen sich neben den Futter- und Körnerleguminosen als Hauptkulturen am besten Untersaaten im Getreide mit Weiß- und Inkarnatklee (Trifolium incarnatum L.). Diese beiden Arten blühen, wenn der Raps und andere Massentrachten (z. B. Obst) bereits verblüht sind (Ministerium für Ländlichen Raum und Verbraucherschutz, 2019). Beyer et al. (2019) zeigten, dass Hummeln mit langen Zungen häufiger in Landschaften mit Ackerbohnen vorkommen als in vergleichbaren Landschaften ohne Ackerbohnen. Dieser Effekt blieb auch nach der Blüte der Ackerbohnen erhalten. Bei der Züchtung von Körnerleguminosen wird bislang neben dem Ertrag die Fähigkeit zur Selbstbestäubung der Pflanzen und nicht das Pollen- und Nektarangebot sowie eine angepasste Blütenform gefördert (Palmer et al., 2009), obwohl Studien gezeigt haben, dass die Bestäubung durch Insekten sich sogar bei weitestgehend selbstbestäubenden Arten wie Sojabohne oder Erbse positiv auf den Ertrag auswirken (Chiari et al., 2005, Monasterolo et al., 2015, Naeem et al., 2018). Auf diese Weise bleibt viel Potenzial von Körnerleguminosen zur Schließung von Trachtlücken und damit die Förderung von Ökosystemleistungen durch Bestäuber ungenutzt. Dieser Nachteil müsste gegenüber möglichen ökonomischen Vorteilen der Selbstbestäubung abgewogen werden. Über den floralen Nektar hinaus bieten viele Hülsenfrüchte extrafloralen Nektar, der nicht nur von Bestäubern, sondern auch von anderen nützlichen Arten wie parasitoiden Wespen (Géneau et al., 2012) genutzt werden kann.

Abb. 7. Blütenvielfalt von Körnerleguminosen. Oben: Ackerbohne, Sojabohne, buntblühende Wintererbse, weißblühende Sommererbse; unten: Blaue Lupine, Gelbe Lupine, Saatwicke und Platterbse (jeweils von links nach rechts).
Allgemein wird davon ausgegangen, dass ein durch Leguminosen verbessertes Stickstoff- und Eiweißangebot der ganzen trophischen Kette zu Gute kommt. So profitieren herbivore und omnivore Arthropoden direkt von den stickstoff- und eiweißreichen Ernterückständen der Leguminosen (Cass et al., 2014). Ob dies zu einer einseitigen Förderung von Schädlingen oder auch zur Kontrolle dieser Schädlinge durch deren natürliche Feinde führen kann, ist nicht abschließend geklärt. Nur wenige Einzelstudien sind derzeit hierzu verfügbar. Der Artenreichtum der Laufkäfer (Carabidae) war in Schlägen mit Sojabohnenanbau tendenziell höher als in anderen Ackerfrüchten wie Mais, Hafer, Sonnenblume (Helianthus annuus L.) und Weizen (Ellsbury et al., 1998; Larsen et al., 2003; O'Rourke et al., 2008; de la Fuente et al., 2014; Molina et al., 2014). Young & Edwards (1990) berichteten in einem Review-Artikel über einen signifikant erhöhten Artenreichtum bei Spinnen (Araneae) in Sojabohnen im Vergleich zu vielen anderen Kulturarten wie Reis (Oryza sativa L.), Sorghumhirse [Sorghum bicolor (L.) MOENCH], Mais und Zuckerrohr (Saccharum officinarum L.). Einzig bei anderen Leguminosen (insbesondere mehrjährigen) zeigte sich ein höherer Artenreichtum und eine höhere Aktivität als bei Sojabohnen. Diese Beziehung wurde in einer wissenschaftlichen Studie näher untersucht, in der die Luzerne als ein- oder mehrjährige Kultur mit der einjährigen Sojabohne verglichen wurde. Es zeigten sich in Luzerne ab dem zweiten Jahr stets eine höhere Aktivität und ein gesteigerter Artenreichtum bei Spinnen als in Sojabohnen (Culin & Yeargan, 1983a und 1983b). Cass et al. (2014) führten in einem Bericht zum Einfluss von Leguminosen auf die Biodiversität zahlreiche Quellen auf, die den positiven Einfluss des Leguminosenanbaus (besonders Sojabohne, Luzerne, Lupine und Klee) auf die Aktivität räuberischer Arthropoden sowie auf parasitoide Wespen zeigten (Curry, 1986; Osler et al., 2000; Hooks & Johnson, 2001; Midega et al., 2009). Der förderliche Effekt von Sojabohnen auf räuberische Arthropoden konnte bis in die Folgekultur hinein nachgewiesen werden (Brust et al., 1986). Neben solchen Vorfruchteffekten können auch Nachbarschaftseffekte zum Tragen kommen. Schulz et al. (2019) zeigten, dass die Dichte von Blattlausmumien in Weizenfeldern, die an Ackerbohnenfelder angrenzten, höher war als in Feldern, die neben anderen Weizenfeldern lagen. Die Dichte der Blattlausmumien weist auf die Parasitierungsrate der Blattläuse hin. Gleichzeitig wurde die Dichte der herbivoren Insekten durch die Nachbarschaft zur Ackerbohne nicht erhöht.
Regenwürmer (Lumbricidae) werden aufgrund ihrer essentiellen Rolle in Agrarökosystemen als Indikatoren für Biodiversität und Bodenfruchtbarkeit verwendet (Bartz et al., 2013). Leguminosen beeinflussen sowohl den Artenreichtum als auch die Aktivität von Regenwürmern. Der Anbau von Futterleguminosen wie z. B. Klee (Abb. 8) hatte sowohl im Gemisch im Grünland als auch im Zwischenfruchtanbau einen positiven Effekt auf die Aktivität und den Artenreichtum von Regenwürmern (Schmidt et al., 2003; Jordan et al., 2004; Birkhofer et al., 2011). Bei den Körnerleguminosen ergibt sich aus der Literatur jedoch kein klares Bild. Smith et al. (2005) berichteten von höheren Regenwurmaktivitäten in Sojabohnen verglichen mit anderen Ackerkulturen. Ashworth et al. (2017) wiederum präsentierten widersprüchliche Ergebnisse. In einem Feldexperiment fand sich in Sojabohnen eine höhere Aktivitätsdichte als im Mais. In einem zweiten Feldexperiment hingegen förderte Mais die Regenwurmpopulation stärker als Sojabohne.

Abb. 8. Weißklee (Trifolium repens L.) zur Futternutzung.
Generell kann aus der bestehenden begrenzten Kenntnislage geschlussfolgert werden, dass die Integration von Leguminosen in Fruchtfolgen die Biodiversität und die mit ihr verbundenen Ökosystemleistungen potenziell fördern kann. Eine positive Wirkung hängt jedoch auch von der Art und Intensität der Bewirtschaftung der Kulturen in den Fruchtfolgen ab. Fruchtfolgen mit Leguminosen sind im Hinblick auf Biodiversität und Ökosystemleistungen noch sehr wenig erforscht. Dadurch fehlt es an Wissen über den Einfluss von Leguminosen unter spezifischen Standortbedingungen (z. B. Niederschlagsmenge, Lufttemperatur, Bodenwassergehalt, Bodenart) im Zusammenwirken mit pflanzenbaulichen Maßnahmen auf verschiedene Organismengruppen. Für keine der Leguminosenarten in Fruchtfolgen liegen bislang ausreichend belastbare Kenntnisse vor, um gesicherte Aussagen über deren Einfluss auf die Biodiversität und Ökosystemleistungen in mitteleuropäischen Ackerbausystemen treffen zu können.
Die Treibhausgasbilanz im Ackerbau wird überwiegend vom Input an organischem Kohlenstoff und Stickstoff sowie synthetischem Stickstoff bestimmt. Es gibt dabei vier wesentliche Pfade, durch die Treibhausgasemissionen freigesetzt werden können:
1. Natürlich ablaufende Prozesse des Stickstoffkreislaufs, d. h. Nitrifikation, Denitrifikation und andere Prozesse (Butterbach-Bahl et al., 2013) produzieren aus Düngerstickstoff Lachgas (N2O) als Zwischen- und/oder Endprodukt, welches dann freigesetzt wird. Diese Prozesse finden entweder direkt auf dem Feld (direkte Emissionen) oder nach der Nitratauswaschung oder Ammoniakdeposition an anderer Stelle (indirekte Emissionen) statt. |
2. Bodenkohlenstoff wird in wasserungesättigten Böden mineralisiert und Menge und Qualität des Inputs von Kohlenstoff in den Boden (durch organische Düngung und ober- und unterirdische Ernteresiduen) zusammen mit Klima- und Bodeneigenschaften (die die Mineralisierungsrate beeinflussen) bestimmen, ob es netto zu einem Verlust oder einer Sequestrierung von Kohlenstoff kommt. |
3. Die Produktion synthetischer Düngemittel erzeugt Treibhausgasemissionen durch den Energieverbrauch und Lachgasemissionen während der Herstellung. |
4. Der Energieverbrauch der Landmaschinen führt zu CO2-Emissionen. |
Gemäß IPCC-Regularien (IPCC, 2006) wird nur Punkt 1 dem Berichtssektor Landwirtschaft zugeordnet. Punkt 2 wird im Sektor Landnutzung, Landnutzungsänderungen und Forstwirtschaft (LULUCF) berichtet, während die Punkte 3 und 4 in den Sektoren Industrie und Energie berichtet werden. Dennoch sind Einsparungen in allen diesen Bereichen notwendig (BMU, 2016). Insgesamt setzt das Klimaschutzgesetz (Bundesgesetzblatt, 2019) für die Landwirtschaft (einschließlich Energieverbrauch) das ambitionierte Ziel einer Treibhausgaseinsparung von 34 % gegenüber 1990.
Eine Verstärkung des Leguminosenanteils in Fruchtfolgen ist allgemein als Klimaschutzmaßnahme anerkannt (vgl. Newell Price et al., 2011). Die Wirkung entsteht durch die Substitution synthetischer Stickstoffdünger durch die biologische Stickstofffixierung (Wang et al., 2018). Außerdem werden Überfahrten zum Zwecke der Düngung vermieden und so Energie eingespart. Da die biologische Stickstofffixierung eine wesentliche Grundlage des ökologischen Landbaus darstellt, ist auch das Ziel, mehr Fläche ökologisch zu bewirtschaften (Bundesregierung, 2017), ein Beitrag zum Klimaschutz.
Rees et al. (2013) quantifizieren für Großbritannien jährliche Einsparpotentiale zwischen 0,5 und 1 t CO2-Äquivalente pro Hektar durch verstärkte Nutzung der Stickstofffixierung von Klee und Einführung zusätzlicher Fruchtfolgefelder (einschließlich Leguminosen) in die Fruchtfolgen. Allerdings ist das Einsparpotential stark von Leguminosenart und Klima abhängig, einerseits aufgrund des Einflusses dieser Faktoren auf die Stickstofffixierungsleistung (Liu et al., 2011) und anderseits wegen der starken Abhängigkeit der Lachgasemissionen von Bodenfeuchte und –temperatur (Butterbach-Bahl et al., 2013) zu Zeiten hoher Stickstoffverfügbarkeit in der Fruchtfolge (z. B. nach der Ernte bzw. Einarbeitung von Leguminosen). Zudem ist die aus Ernteresiduen entstehende Lachgasmenge noch unklar. Das IPCC (2006) traf hier die grobe Annahme, dass für Stickstoff aus Ernteresiduen derselbe Emissionsfaktor (1 % pro kg zugeführtem N) gilt wie für Düngerstickstoff, dass also unabhängig von der Art des Stickstoffs (mineralisch, organisch aus Wirtschaftsdüngern oder Pflanzenresiduen) 1 % des Stickstoffinputs als Lachgas emittiert wird. Zukünftig soll angenommen werden, dass (außerhalb von Trockenregionen) die direkten Lachgasemissionen aus dem Stickstoff der Ernteresiduen 37,5 % der Emissionen nach Applikation synthetischer Dünger betragen (IPCC, 2019). Die Basis hierfür bildet jedoch eine rein empirische Metaanalyse ohne Berücksichtigung von Prozesszusammenhängen (IPCC, 2019, Annex 11A.2) und keine gezielten Studien; hier besteht also noch erheblicher Forschungsbedarf.
Es wird derzeit davon ausgegangen, dass der biologische Prozess der Stickstofffixierung selbst kein Lachgas erzeugt (IPCC, 2006; Zhong et al., 2009). Insgesamt ist aber noch nicht geklärt, ob ein gegenüber dem Status quo verstärkter Leguminosenanbau zu erhöhten Lachgasemissionen auf dem Feld führen könnte. Basche et al. (2014) stellten in ihrer Metaanalyse fest, dass sich die Lachgasemissionen durch Integration von legumen Zwischenfrüchten in Fruchtfolgen erhöhen. Allerdings schlussfolgerten sie auch, dass mehr ganzjährige Messungen erforderlich sind, da kurzfristige Effekte zum Teil später in der Folgefrucht wieder ausgeglichen werden.
Bisher gibt es nur wenige Erkenntnisse zur Wirkung des Leguminosenanbaus auf die Bodenkohlenstoffvorräte. Die Forschung auf diesem Gebiet ist langwierig, weil die geringen relativen Veränderungen nur über Zeiträume von mehr als 10 Jahren zweifelsfrei bestimmt werden können. Der Anbau von Körnerleguminosen führt infolge des im Vergleich zu Getreidearten geringeren C-Inputs eher zum Verlust von Bodenkohlenstoff (Plaza-Bonilla et al., 2016). Poeplau & Don (2015) konnten in ihrer Metaanalyse keinen signifikanten Unterschied in der Wirkung von Zwischenfrüchten mit und ohne Leguminosen auf den Vorrat von Bodenkohlenstoff feststellen, belegten aber, dass Zwischenfrüchte allgemein positiv auf den Kohlenstoffvorrat wirken. Dieser Effekt kann die negative Wirkung der Körnerleguminosen auf den Kohlenstoffvorrat im Boden ausgleichen (Plaza-Bonilla et al., 2016).
Angesichts des Klimawandels werden zukünftig auch Fragen der Auswirkungen von Klimaveränderungen auf Treibhausgasemissionen an Bedeutung gewinnen. So zeigten beispielsweise Lam et al. (2012), dass der Anstieg der atmosphärischen CO2-Konzentration zu einer stärkeren Fixierungsleistung von Leguminosen führt, weil sowohl Anzahl und Masse der Wurzelknöllchen als auch die Nitrogenase-Aktivität zunehmen. Dies würde einerseits die Düngersubstitutionsleistung verstärken, könnte aber auch zur Erhöhung von direkten und indirekten Lachgasemissionen und zur N-Auswaschung aus den Residuen führen. Die Forschung sollte daher zunehmend den Blick auch auf die Interaktion von Klimaschutz und Klimaanpassung richten.
Da kleinkörnige Leguminosen im konventionellen Ackerbau kaum eine Rolle spielen und in der Regel nicht als Hauptfrucht angebaut werden, existieren auch kaum Untersuchungen zur Wirtschaftlichkeit ihres Anbaus. Deshalb wird nachfolgend der Wissensstand zur Wirtschaftlichkeit des Anbaus großkörniger Leguminosen im konventionellen Ackerbau zusammengefasst. Weiterhin weisen einige Autoren auf die Bedeutung der Vermarktungsstrukturen für die Wirtschaftlichkeit des Leguminosenanbaus hin (Kezeya Sepngang et al., 2018; Preissel et al., 2017). Allerdings finden sich, mit Ausnahme von Analysen des Futterwertes, keine systematischen Analysen zu diesem Themenbereich, so dass sich die nachfolgende Literaturauswertung auf die innerbetriebliche Wettbewerbsfähigkeit auf Ebene des landwirtschaftlichen Betriebs beschränkt.
Preissel et al. (2015) haben in einer Metastudie Arbeiten aus mehreren europäischen Ländern genutzt, um Deckungsbeiträge von Leguminosen mit denen von Getreide und Raps zu vergleichen. Dabei zeigte sich, dass Leguminosen oftmals geringere Deckungsbeiträge aufweisen als die Nicht-Leguminosen. Die Deckungsbeitragsdefizite lagen bei maximal 580 € ha–1.
Sojabohnen zeigten hingegen in einigen Studien Wettbewerbsvorteile gegenüber Getreide. In Rheinland-Pfalz (Riedesser, 2012) und Südfrankreich (Mahmood, 2011) lagen die Deckungsbeiträge von Sojabohnen mit bis zu 200 € ha–1 über denen von Getreide. Untersuchungen der Landesanstalt für Landwirtschaft Bayern (LfL) zeigten, dass Sojabohnen (bei Vertragsanbau) wettbewerbsfähig zu Weizen, Körnermais und Winterraps angebaut werden können und um bis zu 600 € ha–1 höhere Deckungsbeiträge erzielen als Ackerbohnen und Blaue Lupine (Lupinus angustifolius L.). Grund für die höhere Wettbewerbsfähigkeit der Sojabohnen gegenüber anderen großkörnigen Leguminosen ist ihr etwa doppelt so hoher Marktpreis. Entsprechende Untersuchungen der Gesamtfruchtfolgen mit Winterweizen, Stoppelweizen und Winterraps zeigten, dass der Gesamtdeckungsbeitrag um etwa 20 € ha–1 steigt, wenn Sojabohnen in die Fruchtfolge integriert werden (LfL, 2015).
Bei Erbsen und Ackerbohnen wurden lediglich in Einzelstudien aus Polen (LMC international, 2009), Deutschland (Zilles, 2010) und Finnland (Peltonen-Sainio & Niemi, 2012) höhere Deckungsbeiträge als für Getreide ausgewiesen. Die Mehrzahl der Studien wies erhebliche Deckungsbeitragsdefizite von 300 bis 500 € ha–1 gegenüber Getreide auf. Einschränkend zu den bisherigen Arbeiten ist anzumerken, dass als Vergleichskulturen relativ wettbewerbsstarke Fruchtarten wie Weizen und Mais unterstellt wurden. Leguminosen müssen in der Regel jedoch mit den schwächeren Fruchtfolgefeldern wie Stoppelweizen konkurrieren. Neben den geringeren Deckungsbeiträgen ergeben sich aufgrund der Ertragsschwankungen in der Regel auch deutlich höhere Volatilitäten in den Deckungsbeiträgen für Leguminosen. Aus diesem Grund sehen von Richthofen et al. (2006b) vor allem pflanzenbauliche Vorteile als Argument für den Anbau von Leguminosen.
Preissel et al. (2017) haben Modellkalkulationen zur Wirtschaftlichkeit des Leguminosenanbaus mit und ohne Berücksichtigung von Vorfruchtwerten für fünf Regionen in Europa (Brandenburg, Kalabrien, Ost-Schottland, West-Schweden, Süd-Rumänien) durchgeführt. Ohne Berücksichtigung von Vorfruchtwerten waren die Deckungsbeiträge von Leguminosen gegenüber den Alternativkulturen nur an den Standorten in Schweden, England und Rumänien wettbewerbsfähig. Für Brandenburg wurde ein Deckungsbeitragsdefizit von 300 bis 320 € ha–1 angegeben. Auch hier wurden als Hauptursache für die Wettbewerbsnachteile die Mindererträge von Leguminosen gegenüber den Alternativkulturen genannt, die in der Regel nicht durch höhere Preise ausgeglichen werden können. Unter Berücksichtigung des Vorfruchtwertes stieg die Wettbewerbsfähigkeit der Leguminosen um 120–300 € ha–1 an.
Insgesamt ist festzustellen, dass die Wettbewerbsfähigkeit von Leguminosen häufig nicht gegeben ist. Einige Studien zeigen jedoch, dass vor allem der Sojabohnenanbau unter Berücksichtigung des Vorfruchtwertes wettbewerbsfähig sein kann. Der Vorfruchtwert bestimmt sich durch die Vorteile für die Folgefrüchte in Form eines höheren Ertrages und verringerter Kosten im Bereich Düngung, Pflanzenschutz und Bodenbearbeitung (Weitbrecht & Pahl, 2000; Schäfer, 2013). Aus diesem Grund beschäftigen sich viele wissenschaftliche Arbeiten im Bereich Ökonomik von Leguminosen mit dem Vorfruchtwert. Nachfolgend werden die bisherigen Ergebnisse dargestellt.
In Tab. 2 sind die bisherigen Arbeiten zum monetären Vorfruchtwert zusammengefasst. Daraus ist zunächst zu entnehmen, dass die Vorfruchtwerte zwischen den verschiedenen Studien mit einer Spannweite von 100 bis 500 € ha–1 stark variieren. Hauptursache dieser erheblichen Spannweite sind vor allem Unterschiede in den Annahmen, die nachfolgend beschrieben werden:
1. Mehrertrag der Folgekultur: Die meisten Studien wiesen Mehrerträge der Folgekultur im Vergleich zur Vorfrucht Weizen von 0 bis 15 dt ha–1 aus (Albrecht & Guddat, 2004; Alpmann & Schäfer, 2014; Zerhusen-Blecher et al., 2018). Lediglich bei Lütke-Entrup et al. (2005) ergaben sich an einzelnen Versuchsstandorten mit bis zu 25 dt ha–1 deutlich höhere Ertragsvorteile für die Folgekultur. Einige Autoren wiesen auch geringe Mehrerträge der zweiten Folgekultur im Bereich von 1 bis 2 dt ha–1 aus (Albrecht & Guddat, 2004; Alpmann et al., 2013). |
2. Preisannahmen zu den Folgekulturen: Erhebliche Unterschiede hinsichtlich des monetären Vorfruchtwertes ergeben sich aus völlig unterschiedlichen Preisannahmen. Hier reichen die Spannweiten hinsichtlich des Getreidepreisniveaus von 100 € t–1 für Gerste bei Albrecht & Guddat (2004) bis 250 € t–1 für Weizen bei Alpmann et al. (2013). |
3. N-Einsparung der Folgekultur: In den meisten Studien wurde eine N-Nachlieferung für die Folgekultur von 0 bis 25 kg N ha–1 unterstellt (Albrecht & Guddat, 2004; Alpmann & Schäfer, 2014; Zerhusen-Blecher et al., 2018). Auch die Preisannahmen zur monetären Bewertung unterschieden sich teilweise erheblich und reichten von 0,5 € pro kg Stickstoff (Albrecht & Guddat, 2004) bis 1 € pro kg Stickstoff (Alpmann & Schäfer, 2014). |
4. Einsparung bei Pflanzenschutzmitteln in der Folgekultur: Die meisten Autoren unterstellten keine eingesparten Pflanzenschutzkosten für die Folgekultur. Lediglich von Richthofen et al. (2006a) sowie Alpmann & Schäfer (2014) berücksichtigten Einsparungen beim Fungizideinsatz in der Folgekultur von bis zu 50 € ha–1, in der Getreide als Referenzfrucht eingesetzt wurde. |
5. Einsparung bei der Bodenbearbeitung: Lediglich in vier Studien wurden Einsparpotentiale in der Bodenbearbeitung der Folgekultur aufgrund einer verbesserten Bodenstruktur/Bodengare berücksichtigt (Schäfer & Lütke-Entrup, 2009; Alpmann et al., 2013; Preissel et al., 2015; Zerhusen-Blecher et al., 2018). Die absolute Höhe des Einsparpotentials reichte von 23 € ha–1 (Zerhusen-Blecher et al., 2018) bis 125 € ha–1 (Lütke-Entrup et al., 2005). |
Tab. 2. Bisherige Untersuchungen zum Vorfruchtwert von Leguminosen.
Studie | Mehrertrag | Einsparung | Einsparung | Einsparung | Vorfruchtwert |
9,2 | 5 – 24 | k.A. | k.A. | 118 – 138 | |
0,5 – 25 | 0–20 | 5 – 24 | k.A. | 98 – 380 | |
0,5 – 1 | 30 | 35 | k.A. | 152 – 204 | |
6,3 | 27 | k.A. | 35 | > 244 | |
5 – 15 | 5 – 35 | 0 – 50 | 20 – 60 | 127 – 471 | |
5 – 15 | 23 – 31 | < 50 | 70 – 125 | 160 – 300 | |
6,6 – 7,5a | 28 – 32 | k.A. | 23 – 42 | 155 – 188 | |
a) Der Mehrertrag ist in Getreideeinheiten angegeben. | |||||
Insgesamt zeigen die bisherigen Studien erhebliche Schwankungsbreiten in der Wettbewerbsfähigkeit. Dass dies häufig auch durch regionale Unterschiede hervorgerufen wird, bestätigt ein Blick in die deutsche Regionalstatistik. Auf Ebene der Landkreise schwanken die Ertragsrelationen von Weizen zu Ackerbohne im Mittel der Jahre 2012 – 2016 zwischen 1: 0,3 und 1: 0,7. Unter der Annahme, dass das Verhältnis der Kosten zwischen den Standorten in etwa konstant bleibt, ist davon auszugehen, dass die Wettbewerbsfähigkeit regional sehr unterschiedlich ausfällt.
Landwirte sind trotz ökonomisch günstiger Vorfruchtwirkungen und vorteilhafter ökologischer Aspekte gegenüber einem verstärkten Anbau von Leguminosen nur dann aufgeschlossen, wenn dieser wirtschaftlich attraktiv ist. Deshalb sollte es künftig vordringlich darum gehen, dabei zu helfen, die bestehenden Hemmnisse für eine Anbauausweitung zu überwinden. Nachhaltige Anbausysteme mit Leguminosen erfordern neben pflanzenbaulichen Optimierungsstrategien insbesondere auch die Berücksichtigung phytopathologischer Aspekte sowie die Züchtung und Bereitstellung resistenter Sorten.
Die Ausweitung des Leguminosenanbaus kann nur gelingen, wenn neu geschaffenes Wissen in die landwirtschaftliche Praxis transferiert und dort entsprechend umgesetzt wird. Hierzu müssen Ausbildung und Weiterbildung sowie die Beratung auf allen Ebenen des Agrarsektors intensiviert werden. Dabei kommt den Demonstrationsnetzwerken Sojabohne, Erbse/Bohne, Lupine und Feinsamige Leguminosen eine wichtige Bedeutung bei der Wahrnehmung folgender Aufgaben zu: (a) Betreuung der Internetplattformen, (b) Informations- und Kontaktbörse, (c) Anbauberatung, (d) Mitwirkung beim Aufbau von Wertschöpfungsketten, (e) Koordinierung von Leuchtturmprojekten sowie (f) Erkennen und Aufzeigen von Forschungsbedarf.
Künftig sollten vorrangig Projekte initiiert und finanziert werden, die mit hoher Wahrscheinlichkeit zu einer deutlichen Ausweitung des Leguminosenanbaus beitragen können. Notwendig ist auch die Anpassung der Bearbeitungsdauer an die jeweilige Fragestellung. So sind zur Erzielung substanzieller Ergebnisse im Bereich der Fruchtfolgeforschung Projektlaufzeiten von deutlich mehr als drei Jahren erforderlich. Neue Projekte sollten vorzugsweise im Rahmen von interdisziplinären Forschungsverbünden und im engen Austausch mit den jeweiligen Demonstrationsnetzwerken realisiert werden. So könnten beispielsweise längerfristige transdisziplinäre Projekte zur Fruchtfolgeforschung an verschiedenen Standorten Deutschlands etabliert werden und mit on-farm-Forschungsansätzen auf Betrieben der Demonstrations-Netzwerke oder in „Landschaftslaboren“ mit Betriebsverbünden unter Beteiligung der ökonomischen Forschung sowie unter Berücksichtigung von Treibhausgasemissionen und Biodiversitätsmaßnahmen verknüpft und wissenschaftlich begleitet werden.
Der vorrangige Forschungsbedarf in den Themenbereichen Pflanzenbau, Pflanzenschutz, Biodiversität, Klimaschutz und Ökonomie ist in Tab. 3 aufgelistet.
Tab. 3. Forschungsbedarf zu Wirkungen der Integration von Leguminosen in Fruchtfolgen.
Themenbereich | Rahmenbedingungen | Vordringlicher Forschungsbedarf |
Pflanzenbau | Viele wichtige pflanzenbauliche Fragen zum Anbau von Körnerleguminosen und deren Integration in Fruchtfolgen wurden bereits im Rahmen von früheren Forschungsprojekten beantwortet. Insbesondere ist die Größenordnung der Vorfruchtwirkung von Leguminosen hinlänglich beschrieben. Forschungslücken bestehen jedoch weiterhin im Bereich züchterischer Verbesserungen, Optimierung von Anbauverfahren und Entwicklung neuer Nahrungs- und Futtermittel. | Verbesserung der abiotischen und biotischen Stresstoleranz (z. B. Kühletoleranz bei Sojabohnen, Winterhärte bei Winterformen der Ackerbohnen und Erbse) |
Erhöhung der Ertragssicherheit von Leguminosen durch züchterische Verbesserung der Krankheits- und Schädlingsresistenz sowie der Unkrautunterdrückung | ||
Minderung bzw. Eliminierung antinutritiver und anderer wertmindernder Inhaltsstoffe | ||
Ermittlung der innerartlichen Variabilität für die Stickstofffixierleistung | ||
Förderung von Maßnahmen und Verfahren zur Bekämpfung samenbürtiger Schädlinge bei großkörnigen Leguminosen | ||
Bearbeitung des Themenkomplexes „Leguminosenmüdigkeit“ in Fruchtfolgen (z. B. mögliche Anbauhäufigkeit von Leguminosen, Wechselwirkungen zwischen verschiedenen Leguminosenarten und mit anderen Kulturarten) | ||
Bewertung legumer Zwischenfrüchte bzw. Zwischenfruchtgemenge hinsichtlich Vorfruchtwert und Ökosystemleistungen (z. B. Stickstofffixierung, Stickstoffverwertung und Stickstoffauswaschung) | ||
Entwicklung und Erprobung von innovativen Anbauverfahren für Körner- und Futterleguminosen (z. B. Fruchtfolge- und Anbaumanagement sowie Integration und Weiterentwicklung von Gemengeanbausystemen) | ||
Förderung der Entwicklung wirtschaftlich erfolgversprechender Nahrungs- und Futtermittelprodukte auf Basis von Leguminosen | ||
Pflanzenschutz | Bei einer Erhöhung des Leguminosenanteils in der Fruchtfolge ist grundsätzlich mit einem vermehrten Auftreten von leguminosenspezifischen Schadorganismen zu rechnen. Verschärft wird dieses Problem durch die abnehmende Verfügbarkeit chemischer Pflanzenschutzmittel. Durch Konzentration auf einige wenige Wirkstoffe zur Bekämpfung von Schadorganismen und Unkräutern ist zudem mit verstärkten Resistenzbildungen zu rechnen. | Entwicklung von Verfahren für die Anwendung von Saatgutbehandlungsmitteln zur Kontrolle bodenbürtiger Schadpilze (z. B. Pythium spp., Rhizoctonia spp., Aphanomyces spp., Sclerotinia spp.) und Nematoden (z. B. Meloidogyne spp., Pratylenchus spp., Heterodera spp.) |
Entwicklung und Erprobung wirksamer Maßnahmen und Verfahren zur Kontrolle von tierischen Schaderregern wie z. B. Erbsenwickler (Cydia nigricana), Erbsengallmücke (Contarinia pisi), Ackerbohnen- und Erbsenkäfer (Bruchus rufimanus, Bruchus pisorum) und Blattrandkäfern (Sitona spp.) | ||
Entwicklung und Nutzung nichtchemischer Verfahren zur Regulierung und Bekämpfung von pilzlichen Schaderregern (z. B. Ascochyta spp., Uromyces viciae-fabae, Peronospora spp., Botrytis spp. und Colletotrichum spp.) | ||
Entwicklung von Prognoseverfahren und Entscheidungshilfen für die Anwendung von Pflanzenschutzmitteln auf der Grundlage des Schadschwellenprinzips | ||
Biodiversität | Zur Auswirkung der Integration von Leguminosen in Fruchtfolgen auf die Biodiversität besteht generell noch Forschungsbedarf. Ob und wenn ja, welche Organismengruppen von Leguminosen gefördert werden ist nicht hinreichend untersucht. Daher existieren auch kaum Erkenntnisse zu Ökosystemleistungen dieser Organismen im Leguminosenanbau selbst oder in der umgebenden Agrarlandschaft. | Züchtung der Leguminosen auf „Bestäuberfreundlichkeit“ und Bewertung existierender Sorten im Anbausystem hinsichtlich der Bereitstellung von Nahrungsressourcen für Bestäuber (d. h. kommen die Kulturen im Anbau zur Blüte und wenn ja, findet die Ernte während der Hauptblütezeit statt?) |
Katalogisierung der mit den jeweiligen Ackerkulturen sowie Anbausystemen mit und ohne Leguminosen vergesellschafteten Organismengruppen (trophische Gruppen und Taxa) | ||
Agrarökosystemare Studien zur Wirkung von Leguminosen in Anbausystemen auf die Biodiversität von der Feld- bis zur Landschaftsskala | ||
Vorfrucht- und Nachbarschaftseffekt von Leguminosen auf Herbivoren und Prädatoren | ||
Quantifizierung der Ökosystemdienstleistungen (insbesondere natürliche Schädlingskontrolle, Bestäubung, Bodenfruchtbarkeit) durch Leguminosenanbau über mehrere räumliche Skalen (Feld, Betrieb, Landschaft) | ||
Agronomische und sozioökonomische Inwertsetzung der von Leguminosen erbrachten Ökosystemdienstleistungen | ||
Klimaschutz | Bisherige Forschung zu Treibhausgasemissionen im Ackerbau fokussiert häufig stark auf einzelne Fruchtfolgeglieder. Leguminosen wirken sich jedoch auf die gesamte Fruchtfolge aus; dies ist ja gerade das stärkste Argument für ihren Anbau. Es besteht erheblicher Forschungsbedarf bezüglich der Wirkung der Integration von Leguminosen in Fruchtfolgen auf deren gesamte Treibhausgasbilanz. Eine noch besser optimierte Berücksichtigung von Vorfruchtwerten und Fruchtfolgeeffekten gerade bei der Düngeplanung bietet Potenzial zum Klimaschutz. | Auswirkung der Integration von Leguminosen in Fruchtfolgen auf den Bodenkohlenstoffvorrat |
Lachgasemissionen und Nitratauswaschung aus Leguminosenstickstoff beim Stickstofftransfer in die erste und zweite Folgefrucht | ||
Optimierungsmöglichkeiten des Stickstofftransfers in die Folgefrüchte und damit zur Einsparung von Mineraldünger | ||
Entwicklung von praxistauglichen Ansätzen zur optimierten Berücksichtigung der Einsparpotenziale von Stickstoffdüngern in den Folgekulturen von Leguminosen | ||
Ökonomie | Aus Sicht der Ökonomie ist für die Gewinnung aussagekräftiger und vertrauenswürdiger Ergebnisse wichtig, dass die betriebswirtschaftliche Bewertung nach einheitlichen Standards erfolgt. | Systematische Erfassung von Fruchtfolgeeffekten in Praxisbetrieben |
Generierung von Informationen zur Wirtschaftlichkeit des Leguminosenanbaus unter Praxisbedingungen (z. B. durch Netzwerke aus Pilotbetrieben) |
Trotz vielfältiger den Leguminosen zugesprochener und nachgewiesener positiver pflanzenbaulicher und agrarökologischer Wirkungen ist ihr Anteil an der Ackerfläche in Deutschland noch gering. Als Gründe dafür werden oftmals eine unzureichende Ertragssicherheit, Indikationslücken beim Pflanzenschutz, die Leguminosenmüdigkeit bei einem höheren Anteil in Fruchtfolgen und die fehlende Wirtschaftlichkeit angeführt. Für eine umfassende Bewertung der Vor- und Nachteile der Integration von Leguminosen in Fruchtfolgen fehlen jedoch durch entsprechende Feldversuche abgesicherte Ergebnisse zu (1) Saatgutbehandlungs- und Pflanzenschutzmaßnahmen ebenso wie eine mögliche Reduktion von Pflanzenschutzanwendungen in den Folgekulturen, (2) Auswirkungen auf die Biodiversität sowie agronomische und sozio-ökonomische Bewertungen der von Leguminosen erbrachten Ökosystemdienstleistungen, und (3) Effekten auf Bodenkohlenstoffvorrat, Lachgasemissionen und Nitratauswaschung in der gesamten Fruchtfolge. Erst wenn diese Fragen geklärt sind, ist eine Ausdehnung der Leguminosenanbaufläche und eine sichere ökonomische Bewertung der Integration von Leguminosen in Fruchtfolgen möglich.
Die Autoren erklären, dass keine Interessenskonflikte vorliegen.
Abbo, S., B.Y. Saranga, B.Z. Peleg, B.Z. Kerem, B.S. Lev-Yadun, B.A. Gopher, 2009: Reconsidering domestication of legumes versus cereals in the ancient Near East. The Quarterly Review of Biology 84, 29-50, DOI: 10.1086/596462.
Aher, G., L.J. Cihacek, K. Cooper, 2017: An evaluation of C and N on fresh and aged crop residue from mixed long-term no-till cropping systems. Journal of Plant Nutrition 40, 177-186, DOI: 10.1080/01904167.2016.1201505.
Albrecht, R., C. Guddat, 2004: Welchen Wert haben Körnerleguminosen in der Fruchtfolge? Access: 18. Juli 2020, URL:https://docplayer.org/62870012-Welchen-wert-haben-koernerleguminosen-in-der-fruchtfolge.html.
Alpmann, D., J. Braun, B.C. Schäfer, 2013: Fruchtfolgen zielgerichtet auflockern. Access: 18. Juli 2020, URL: https://www4.fh-swf.de/media/downloads/forschung/leguan/automatisch_download/02_Fruchtfolgen_zielgerichtet_auflockern_LuF2013.pdf.
Alpmann, S., B.C. Schäfer, 2014: Der Wert von Körnerleguminosen im Betriebssystem. UFOP-Praxisinformation. Union zur Förderung von Öl- und Proteinpflanzen e. V. Access: 18. Juli 2020, URL: https://www.ufop.de/files/9013/9593/2050/RZ_UFOP_1157_Praxis_Koernerleguminosen_web.pdf.
Andert, S., J. Bürger, J.E. Mutz, B. Gerowitt, 2018: Patterns of pre-crop glyphosate use and in-crop selective herbicide intensities in Northern Germany. European Journal of Agronomy 97, 20-27, DOI: 10.1016/j.eja.2018.04.009.
Ashworth, A., F.L. Allen, A.M. Saxton, D.D. Tyler, 2017: Impact of crop rotations and soil amendments on long-term no-tilled soybean yield. Agronomy Journal 109, 938-946, DOI: 10.2134/agronj2016.04.0224.
Aslam, M., I.A. Mahmood, M.B. Peoples, G.D. Schwenke, D.F. Herridge, 2003: Contribution of chickpea nitrogen fixation to increased wheat production and soil organic fertility in rain-fed cropping. Biology and Fertility of Soils 38, 59-64, DOI: 10.1007/s00374-003-0630-5.
Bartz, M.L.C., A. Pasini, G.G. Brown, 2013: Earthworms as soil quality indicators in Brazilian no-tillage systems. Applied Soil Ecology 69, 39-48, DOI: 10.1016/j.apsoil.2013.01.011.
Basche, A.D., F.E. Miguez, T.C. Kaspar, M.J. Castellano, 2014: Do cover crops increase or decrease nitrous oxide emissions? A meta-analysis. Journal of Soil and Water Conservation 69, 471-482, DOI: 10.2489/jswc.69.6.471.
Becker, K., S. Heilmann, A. Riffel, G. Leithold, S.A. Fischinger, 2013: Wirkung einer Schwefel- und Gülledüngung auf den Trockensubstanz- und Stickstoffertrag eines Futterleguminosenbestandes. In: Beiträge zur 12. Wissenschaftstagung Ökologischer Landbau. Neuhoff, D., et al. (Eds.). Bonn, Verlag Dr. Köster, 220-221.
Bedoussac, L., 2009: Analyse du fonctionnement des performances des associations blé dur-pois d’hiver et blé dur-féverole d’hiver pour la conception d’itinéraires techniques adaptés à différents objectifs de production en systèmes bas-intrants. Access: 18. Juli 2020, URL: http://ethesis.inp-toulouse.fr/archive/00000963/01/bedoussac.pdf.
Beyer, N., F. Kirsch, K. Schulz, G. Everwand, S. Mecke, D. Gabriel, J. Dauber, C. Westphal, 2019: The cultivation of faba beans increases bumblebee abundances at landscape scale. In: GfÖ 2019: Science meets practice; 49th Annual Meeting of the Ecological Society of Germany, Austria and Switzerland; University of Münster, Book of abstracts. Berlin, Gesellschaft für Ökologie, 286.
Birkhofer, K., T. Diekötter, S. Boch, M. Fischer, J. Müller, S. Socher, V. Wolters, 2011: Soil fauna feeding activity in temperate grassland soils increases with legume and grass species richness. Soil Biology and Biochemistry 43, 2200-2207, DOI: 10.1016/j.soilbio.2011.07.008.
BMU (Bundesministerium für Umwelt, Naturschutz und nukleare Sicherheit), 2016: Klimaschutzplan 2050. Klimapolitische Grundsätze und Ziele der Bundesregierung, Publikationsnummer 2261. Access: 18. Juli 2020, URL: https://www.bmu.de/download/klimaschutzplan-2050/.
Böhm, H., 2014: Unkrautregulierung durch Fruchtfolge und alternative Managementverfahren. Julius-Kühn-Archiv 443, 24-36.
Böhm, H., 2016: Einfluss einer Schwefeldüngung auf die Ertragsleistung und ausgewählte Inhaltsstoffe von Kleegras im Ökologischen Landbau. Mitteilung der Gesellschaft für Pflanzenbauwissenschaften 28, 74-75.
Böhm, H., U. Ebert, W. Vogt-Kaute, 2013: Körnerleguminosen im Gemenge. In: Körnerleguminosen anbauen und verwerten. Kuratorium für Technik und Bauwesen in der Landwirtschaft e.V. (KTBL, Eds.), Darmstadt, KTBL-Heft 100, 21-23.
Böhm, H., R. Brandhuber, C. Bruns, M. Demmel, M.R. Finckh, J. Fuchs, A. Gronle, O. Hensel, G. Lux, D. Möller, H. Schmidt, K. Schmidtke, A.-K. Spiegel, W. Vogt-Kaute, D. Werren, K.P. Wilbois, M. Wild, D. Wolf, 2014: Körnerleguminosen und Bodenfruchtbarkeit – Strategien für einen erfolgreichen Anbau. Bundesanstalt für Landwirtschaft und Ernährung (Eds.), Bonn, 59 S.
Böttcher, F., M. Schmidt, 2016: Wie durstig sind Zwischenfrüchte. DLG Mitteilungen 7, 12-13.
Braun, J., J. Brunotte, U. Ernst, J.M. Greef, F.F. Gröblinghoff, M. Heckmann, K. Korte, H. Laser, S. Lutke Börding, V. Richter, P. Schattschneider, W. Stauss, T. Weyer, B.C. Schäfer, 2014: Klimaoptimierte Anpassungsstrategien in der Landwirtschaft. Abschlussbericht. Access: 18. Juli 2020, URL: https://service.ble.de/ptdb/index2.php?detail_id=23354&site_key=145&stichw=Klimaoptimierte + Anpassungsstrategien + in + der + Landwirtschaft&zeilenzahl_zaehler=69#newContent.
Bretag, T.W., L.M. Smith, D.J. Ward, 2001: Effect of soil-borne Ascochyta blight fungi on the grain yield of field peas. In: Science and Technology: Delivering Results for Agriculture? - Proceedings of the 10th Australian Agronomy Conference. Rowe, B., D. Donaghy, N. Mendham (Eds.), Hobart, Australia.
Brust, G.E., B.R. Stinner, D.A. McCartney, 1986: Predator activity and predation in corn agroecosystems. Environmental Entomology 15, 1017-1021, DOI: 10.1093/ee/15.5.1017.
Bundesgesetzblatt, 2019: Gesetz zur Einführung eines Bundes-Klimaschutzgesetzes und zur Änderung weiterer Vorschriften. Bundesgesetzblatt Jahrgang 2019 Teil I Nr. 48. Access: 18. Juli 2020, URL: https://www.bgbl.de/xaver/bgbl/start.xav?startbk=Bundesanzeiger_BGBl&start=//*%5b@attr_id=%27bgbl119s2513.pdf%27 %5d#__bgbl__%2F%2F*%5B%40attr_id%3D%27bgbl119s2513.pdf%27 %5D__1593163854993.
Bundesregierung, 2017: Deutsche Nachhaltigkeitsstrategie, Neuauflage 2016. Access: 18. Juli 2020, URL: https://www.bundesregierung.de/breg-de/themen/nachhaltigkeitspolitik/eine-strategie-begleitet-uns/die-deutsche-nachhaltigkeitsstrategie.
Butterbach-Bahl, K., E.M. Baggs, M. Dannenmann, R. Kiese, S. Zechmeister-Boltenstern, 2013: Nitrous oxide emissions from soils: how well do we understand the processes and their controls? Philosophical Transactions of the Royal Society B Biological Sciences 368 (1621), DOI: 10.1098/rstb.2013.0122.
Cass, S., M. Williams, J. Stout, 2014: Biodiversity and ecosystem services in legume-supported cropping. In: Environmental implications of legume cropping. Legume Futures Report 3.7. Williams, M., J. Stout, B. Roth, S. Cass, V. Papa, B. Rees (Eds.), 44-86. Access: 18. Juli 2020, URL: http://www.legumefutures.de/results/environmentaleffects.html.
Castellazzi, M.S., G.A. Wood, P.J. Burgess, J. Morris, K.F. Conrad, J.N. Perry, 2008: A systematic representation of crop rotations. Agricultural Systems 97, 26-33, DOI: 10.1016/j.agsy.2007.10.006.
Cernay, C., D. Makowski, E. Pelzer, 2018: Preceding cultivation of grain legumes increases cereal yields under low nitrogen input conditions. Environmental Chemistry Letters 16, 631–636, DOI: 10.1007/s10311-017-0698-z.
Chiari, W.C., V.D.A.A.D. Toledo, M.C.C. Ruvolo-Takasusuki, A.J.B.D. Oliveira, E.S. Sakaguti, V.M. Attencia, F.M. Costa, M.H. Mitsui, 2005: Pollination of soybean (Glycine max L. Merril) by honeybees (Apis mellifera L.). Brazilian Archives of Biology and Technology 48, 31-36. DOI: 10.1590/S1516-89132005000100005.
Christen, O., 2001: Ertrag, Ertragsstruktur und Ertragsstabilität von Weizen, Gerste und Raps in unterschiedlichen Fruchtfolgen. Pflanzenbauwissenschaften 5, 33-39.
Congreves, A., A. Hayes, E.A. Verhallen, L.L. van Eerd, 2015: Long-term impact of tillage and crop rotation on soil health at four temperate agroecosystems. Soil and Tillage Research 152, 17-28, DOI: 10.1016/j.still.2015.03.012.
CORDIS, 2014: Final Report Summary - LEGUME ROOT IMPACT (Root longevity and the impact on soil N fertility). Access: 18. Juli 2020, URL: https://cordis.europa.eu/project/id/252830/reporting.
CORDIS, 2015: Final Report Summary - PROLEGSO (Promoting legume-non-legume communities through plant and soil traits). Access: 18. Juli 2020, URL: https://cordis.europa.eu/project/id/254533/reporting.
CORDIS, 2020: Translating knowledge for legume-based farming for feed and food systems. Access: 18. Juli 2020, URL: https://cordis.europa.eu/project/rcn/218512/en.
Culin, J.D., K.V. Yeargan, 1983a: Comparative study of spider communities in alfalfa and soybean ecosystems: Foliage-dwelling spiders. Annals of the Entomological Society of America 76, 825-831, DOI: 10.1093/aesa/76.5.825.
Culin, J.D., K.V. Yeargan, 1983b: Comparative study of spider communities in alfalfa and soybean ecosystems: ground-surface spiders. Annals of the Entomological Society of America 76, 832-838, DOI: 10.1093/aesa/76.5.832.
Curry, J.P., 1986: Above-ground arthropod fauna of four Swedish cropping systems and its role in carbon and nitrogen cycling. Journal of Applied Ecology 23, 853-870, DOI: 10.2307/2403939.
DAFA, 2012: Fachforum Leguminosen, Wissenschaft, Wirtschaft, Gesellschaft – Ökosystemleistungen von Leguminosen wettbewerbsfähig machen. Forschungsstrategie der Deutschen Agrarforschungsallianz. Braunschweig, 57 S.
Degner, J., 2013: Fruchtfolgegestaltung in Thüringer Ackerbaubetrieben aus ökonomischer Sicht. Bulletin SGPW-SSA 26, 9-10.
De la Fuente, E.B., S.A. Suárez, A.E. Lenardis, S.L. Poggio, 2014: Intercropping sunflower and soybean in intensive farming systems: evaluating yield advantage and effect on weed and insect assemblages. NJAS-Wageningen Journal of Life Sciences 70, 47-52, DOI: 10.1016/j.njas.2014.05.002.
DemoNetErbseBohne, 2020: Demonstrationsnetzwerk Erbse/Bohne. Access: 18. Juli 2020, URL: https://www.demoneterbo.agrarpraxisforschung.de/index.php?id=165.
De Notaris, C., J. Rasmussen, P. Sörensen, J.E. Olesen, 2018: Nitrogen leaching: a crop rotation perspective on the effect of N surplus, field management and use of catch crops. Agriculture, Ecosystems & Environment 255, 1-11, DOI: 10.1016/j.agee.2017.12.009.
Destatis, 2015: Landwirtschaftliche Bodennutzung und pflanzliche Erzeugung - Fachserie 3 Reihe 3 – 2014. Access: 18. Juli 2020, URL: https://www.destatis.de/DE/Themen/Branchen-Unternehmen/Landwirtschaft-Forstwirtschaft-Fischerei/Publikationen/Bodennutzung/bodennutzung-erzeugung-2030300147005.html.
Destatis, 2019a: Statistisches Jahrbuch 2019 - Kapitel 19 Land- und Forstwirtschaft. Access: 18. Juli 2020, URL: https://www.destatis.de/DE/Themen/Querschnitt/Jahrbuch/jb-land-forstwirtschaft.pdf?__blob=publicationFile.
Destatis, 2019b: Ackerland nach Hauptfruchtgruppen und Fruchtarten. Access: 18. Juli 2020, URL: https://www.destatis.de/DE/Themen/Branchen-Unternehmen/Landwirtschaft-Forstwirtschaft-Fischerei/Feldfruechte-Gruenland/Tabellen/ackerland-hauptfruchtgruppen-fruchtarten.html.
De Visser, C.L.M., R. Schreuder, F. Stoddard, 2014: The EU’s dependency on soya bean import for the animal feed industry and potential for EU produced alternatives. Oilseeds and Fats, Crops and Lipids 21, DOI: 10.1051/ocl/2014021.
Drury, C.F., W.D. Reynolds, C.S. Tan, N.B. Mc Laughlin, X.M. Yang, W. Calder, T.O. Oloya, J.Y. Yang, 2014: Impacts of 49-51 years of fertilization and crop rotation on growing season nitrous oxide emissions, nitrogen uptake and corn yields. Canadian Journal of Soil Science 94, 421-433, DOI: 10.4141/cjss2013-101.
DüV, 2017: Verordnung über die Anwendung von Düngemitteln, Bodenhilfsstoffen, Kultursubstraten und Pflanzenhilfsmitteln nach den Grundsätzen der guten fachlichen Praxis beim Düngen (Düngeverordnung - DüV). Access: 18. Juli 2020, URL: https://www.gesetze-im-internet.de/d_v_2017/D%C3 %BCV.pdf.
Ellsbury, M.M., J.E. Powell, F. Forcella, W.D. Woodson, S.A. Clay, W.E. Riedell, 1998: Diversity and dominant species of ground beetle assemblages (Coleoptera: Carabidae) in crop rotation and chemical input systems for the Northern Great Plains. Annals of the Entomological Society of America, 91, 619-625, DOI: 10.1093/aesa/91.5.619.
Everwand, G., S. Cass, J. Dauber, M. Williams, J. Stout, 2017: Biodiversity and ecosystem services in legume-supported cropping. In: Legumes in Cropping Systems. Murphy-Bokern, D., F. Stoddard, C. Watson (Eds.), Wallingford, UK, CABI, 55-69.
Finckh, M., T. Yli-Mattila, A. Nykänen, P. Kurki, A. Hannukkala, 2015: Organic temperate legume disease management. In: Plant Diseases and Their Management. Finckh, M., A.H.C. Van Bruggen, L. Tamm (Eds.) St. Paul, Minnesota, U.S.A., The American Phytopathological Society (APS Press), 293-310.
Fischinger, S.A., K. Becker, G. Leithold, 2011: Auswirkungen unterschiedlicher S-Versorgungszustände auf den N Flächenertrag eines Luzerne-Kleegrasbestandes. In: Wissenschaftstagung Ökologischer Landbau. Es geht ums Ganze: Forschen im Dialog von Wissenschaft und Praxis. Leithold, G., et al. (Eds.), Berlin, 183-184.
Free, J.B., 1962: The behaviour of honeybees visiting field beans (Vicia faba). The Journal of Animal Ecology 31, 497-502, DOI: 10.2307/2049.
Gallai, N., J.M. Salles, J. Settele, B.E. Vaissière, 2009: Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecological Economics 68, 810-821, DOI: 10.1016/j.ecolecon.2008.06.014.
Gaudin, A.C.M., T.N. Tolhurst, A.P. Ker, K. Janovicek, C. Tortora, R.C. Martin, W. Deen, 2015: Increasing crop diversity mitigates weather variations and improves yield stability. PloS ONE 10(2), DOI: 10.1371/journal.pone.0113261.
Géneau, C.E., F.L. Wäckers, H. Luka, C. Daniel, O. Balmer, 2012: Selective flowers to enhance biological control of cabbage pests by parasitoids. Basic and Applied Ecology 13, 85-93, DOI: 10.1016/j.baae.2011.10.005.
Gerhards, R., M. Sökefeld, W. Kühbauch, 1998: Einsatz der digitalen Bildverarbeitung bei der teilschlagspezifischen Unkrautkontrolle. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz, Sonderheft XVI, 273-280.
Goulson, D., E. Nicholls, C. Botías, E.L. Rotheray, 2015: Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 347, DOI: 10.1126/science.1255957.
Greef, J.M., F. Höppner, A. Bramm, 2004: Wie werden Fruchtfolgen und Produktionsverfahren im Jahr 2025 aussehen? Landbauforschung Völkenrode, Sonderheft 274, 103-108.
Gronle, A., H. Böhm, J. Heß, 2014: Effect of intercropping winter peas of differing leaf type and time of flowering on annual weed infestation in deep and shallow ploughed soils and on pea pests. Landbauforschung – Agriculture and Forestry Research 64, 31-44, DOI: 10.3220/Lbf_2014_31-44.
Gronle, A., J. Heß, H. Böhm, 2015: Effect of intercropping normal-leafed or semi-leafless winter peas and triticale after shallow and deep ploughing on agronomic performance, grain quality and succeeding winter wheat yield. Field Crops Research 180, 80-89, DOI: 10.1016/j.fcr.2015.05.017.
Gruber, H., C. Wegner, 2017: Schwefeldüngung zu Körnererbsen und Blauen Lupinen auf einem Sandstandort. In: Wissenschaftstagung Ökologischer Landbau. Wolfrum, S., et al. (Eds.), Berlin, Verlag Dr. Köster, 22-25.
Hein, W., 2006: Optimierung von Produktionsverfahren bei Ackerbohnen (Vicia faba L.) zur Sicherung der Ertragsstabilität und Qualität des Erntegutes. Abschlussbericht Ackerbohne des Lehr- und Forschungszentrums Landwirtschaft, LFZ Raumberg-Gumpenstein. Access: 18. Juli 2020, URL: https://www.raumberg-gumpenstein.at/cm4/de/component/jdownloads/viewdownload/1615-2326-ackerbohne-verbesserung-d-saatgutwertes/14011-optimierung-von-produktionsverfahren-bei-ackerbohne-vicia-faba-l-zur-sicherung-der-ertragsstabilitaet-und-qualitaet-des-erntegutes.html.
Heinold, M., M. Kremmer, C. Waibel, T. Schäfer, S. Kübler, 2018: Traktor-integrierte aktive Gerätesteuerung für Hochgeschwindigkeitshacken. In: 38. GIL-Jahrestagung, Digitale Marktplätze und Plattformen. Ruckelshausen, A., A. Meyer-Aurich, K. Borchard, C. Hofacker, J.-P. Loy, R. Schwerdtfeger, H.-H.F. Sundermeier, B. Theuvsen (Eds.), Bonn, Gesellschaft für Informatik e.V., 95-98.
Henne, U., M. Landschreiber, C. Schleich-Saidfar, 2018: Entwicklung nachhaltig wirkender Methoden zur Bekämpfung von Acker- Fuchsschwanz (Alopecurus myosuroides Huds.). Julius-Kühn-Archiv 458, 121-131, DOI: 10.5073/jka.2018.458.018.
Heuser, S., M. Demmel, J. Pfeiffer, M. Gandorfer, 2018: Automatisierte mechanische Unkrautregulierung. In: 38. GIL-Jahrestagung, Digitale Marktplätze und Plattformen. Ruckelshausen, A., A. Meyer-Aurich, K. Borchard, C. Hofacker, J.-P. Loy, R. Schwerdtfeger, H.-H.F. Sundermeier, B. Theuvsen (Eds.), Bonn, Gesellschaft für Informatik e.V., 99-102.
Hocking, P.J., 2001: Organic acids exuded from roots in phosphorus uptake and aluminium tolerance of plants in acid soils. Advances in Agronomy 74, 63-97, DOI: 10.1016/s0065-2113(01)74031-x.
Hooks, C.R., M.W. Johnson, 2001: Broccoli growth parameters and level of head infestations in simple and mixed plantings: Impact of increased flora diversification. Annals of Applied Biology 138, 269-280, DOI: 10.1111/j.1744-7348.2001.tb00112.x.
Huusela-Veistola, E., L. Jauhiainen, 2006: Expansion of cropping increases the risk of pea moth (Cydia nigricana; Lep., Tortricidae) infestation. Journal of Applied Entomology 130, 142-149, DOI: 10.1111/j.1439-0418.2006.01047.x.
IPCC, 2006: IPCC Guidelines for National Greenhouse Gas Inventories, Prepared by the National Greenhouse Gas Inventories Programme. Eggleston, H.S., L. Buendia, K. Miwa, T. Ngara, K. Tanabe K (Eds.). Japan, IGES, Access: 18. Juli 2020, URL: https://www.ipcc-nggip.iges.or.jp/public/2006gl/.
IPCC, 2019: 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories. Gómez, D., W. Irving (Eds.). Access: 19. März 2020, URL: https://www.ipcc-nggip.iges.or.jp/public/2019rf/index.html.
Jensen, E.S., 2006: Grain legume functions in crop rotations. In: Grain legumes and the environment: how to assess benefits and impacts. AEP (Eds.), Zurich, AEP and FAL, 49-54.
Jordan, D., R.J. Miles, V.C. Hubbard, T. Lorenz, 2004: Effect of management practices and cropping systems on earthworm abundance and microbial activity in Sanborn Field: a 115-year-old agricultural field. Pedobiologia 48, 99-110, DOI: 10.1016/j.pedobi.2003.06.001.
Kahnt, G., 2008: Leguminosen im konventionellen und ökologischen Landbau. Frankfurt am Main, DLG-Verlag.
Kezeya Sepngang, B., I. Stute, W. Stauss, B.C. Schäfer, M. Mergenthaler, 2018: Möglichkeiten zur Bildung von verwertungsorientierten Preisindikatoren für Futtererbsen und Ackerbohnen im Vergleich zur veröffentlichten Marktpreisberichterstattung. Berichte über Landwirtschaft, Zeitschrift für Agrarpolitik und Landwirtschaft 96. Access: 18. Juli 2020, URL: https://buel.bmel.de/index.php/buel/article/view/226.
Kolbe, H., 2009: Fruchtfolgegrundsätze im ökologischen Landbau. Abteilung Pflanzliche Erzeugung, Sächsisches Landesamt für Umwelt, Landwirtschaft und Geologie. Access: 18. Juli 2020, URL: http://orgprints.org/15100/.
Köpke, U., 2013: Bedeutung der Fruchtfolge aus phytomedizinischer, pflanzenbaulicher und betriebswirtschaftlicher Sicht. Bulletin SGPW-SSA 26, 2-3.
Köpke, U., R. Rauber, K. Schmidtke, H. Goldbach, H.W. Scherer, 2011: Entwicklung neuer Strategien zur Mehrung und optimierten Nutzung der Bodenfruchtbarkeit. BÖLN-Abschlussbericht. Access: 18. Juli 2020, URL: https://www.researchgate.net/publication/277984915_Entwicklung_neuer_Strategien_zur_Mehrung_und_optimierten_Nutzung_der_Bodenfruchtbarkeit.
Köpke, U., R. Rauber, K. Schmidtke, 2016: Optimierung der Unkrautregulation, Schwefel- und Phosphorverfügbarkeit durch Unterfußdüngung bei temporärer Direktsaat von Ackerbohne und Sojabohne. BÖLN-Abschlussbericht. Access: 18. Juli 2020, URL: https://orgprints.org/30761/.
Kraft, J.M., F.L. Pfleger (Eds.), 2001: Compendium of Pea Diseases and Pests. 2nd. Ed., Madison, USA, APS Press.
KTBL, 2013: Körnerleguminosen anbauen und verwerten. Kuratorium für Technik und Bauwesen in der Landwirtschaft e.V. (KTBL, Eds.), Darmstadt, KTBL-Heft 100.
Lam, S.K., D. Chen, R. Norton, R. Armstrong, A.R. Mosier, 2012: Nitrogen dynamics in grain crop and legume pasture systems under elevated atmospheric carbon dioxide concentration: A meta-analysis. Global Change Biology 18, 2853-2859, DOI: 10.1111/j.1365-2486.2012.02758.x.
Larsen, K.J., T.T. Work, F.F. Purrington, 2003: Habitat use patterns by ground beetles (Coleoptera: Carabidae) of northeastern Iowa. Pedobiologia 47, 288-299, DOI: 10.1078/0031-4056-00192.
LfL (Bayerische Landesanstalt für Landwirtschaft), 2015: Wirtschaftlichkeit des Sojabohnenanbaus. Vortrag von Robert Schätzl, Institut für Betriebswirtschaft und Agrarstruktur. Access: 18. Juli 2020, URL: https://www.sojafoerderring.de/wp-content/uploads/2015/04/Forchheim_Wirtschaftlichkeit-Sojabohnen_16.11.2015_Internet.pdf.
Liu, Y.Y., L.H. Wu, J.A. Baddeley, C.A. Watson, 2011: Models of biological nitrogen fixation of legumes. A review. Agronomy for Sustainable Development 31, 155-172, DOI: 10.1051/agro/2010008.
LMC International, 2009: Evaluation of Measures Applied Under the Common Agricultural Policy to the Protein Crop Sector. In: Case Study Monographs. New York, Oxford, Kuala Lumpur, LMC International, Access: 19. März 2020, URL: http://ec.europa.eu/agriculture/eval/reports/protein crops/.
Lütke-Entrup, N., M. Schneider, G. Stemann, F. Gröblinghoff, A. Heißenhuber, H. Pahl, K. Hülsbergen, X. Maidl, C. Sommer, K. Korte, J. Brunotte, H. Kreye, V. Lindwedel, H. Zieseniß, C. Gienapp, R. Schulz, J. Propp, 2005: Bewertung von neuen Systemen der Bodenbewirtschaftung in erweiterten Fruchtfolgen mit Körnerraps und Körnerleguminosen. Access: 18. Juli 2020, URL: https://docplayer.org/12857058-Bewertung-von-neuen-systemen-der-bodenbewirtschaftung-in-erweiterten-fruchtfolgen-mit-koernerraps-und-koernerleguminosen.html.
Magrini, M.B., M. Anton, C. Cholez, G. Corre-Hellou, G. Duc, M.H. Jeuffroy, J.M. Meynard, E. Pelzer, A.S. Voisin, S. Walrand, 2016: Why are grain-legumes rarely present in cropping systems despite their environmental and nutritional benefits? Analyzing lock-in in the French agrifood system. Ecological Economics 126, 152-162, DOI: 10.1016/j.ecolecon.2016.03.024.
Mahmood, F., 2011: Analysis of the conditions for the development of grain legumes in the Midi-Pyrénées region (France), using the APES-FSSIM-Indicators modeling chain. In: Systèmes Intégrés En Biologie, Agronomie, Géosciences, Hydrosciences Et Environnement (SIBAGHE). Montpellier, France, Ecole National Supérieure Agronomique de Montpellier, 168 S.
Männel, M., V. Haberlah-Korr, M. Heupel, 2018: Feldversuche und Monitoring zur Kontrolle von Blattläusen als Virusvektoren an Ackerbohnen und weiteren Leguminosen. 61. Deutsche Pflanzenschutztagung, Universität Hohenheim, Germany, Julius-Kühn-Archiv, 461, 110-111.
Meynard, J.M., A. Messéan, A. Charlier, F. Charrier, M. Farès, M. Le Bail, M.B. Magrini, I. Savini, O. Réchauchère, 2013: Crop diversification: obstacles and levers. Study of farms and supply chains. Synopsis of the study report, INRA. Access: 18. Juli 2020, URL: www6.paris.inra.fr/depe/content/download/3736/35824/version/1/file/Version+Anglaise+Diversification-8pages.pdf.
Midega, C.A., Z.R. Khan, J. Van den Berg, C.K. Ogol, T.J. Bruce, J.A. Pickett, 2009: Non-target effects of the ‘push–pull’ habitat management strategy: parasitoid activity and soil fauna abundance. Crop Protection 28, 1045-1051, DOI: 10.1016/j.cropro.2009.08.005.
Ministerium für Ländlichen Raum und Verbraucherschutz, 2019: Bienenweidekatalog. Verbesserung der Bienenweide und des Artenreichtums. Access: 18. Juli 2020, URL: https://fachdokumente.lubw.baden-wuerttemberg.de/servlet/is/105070/?COMMAND=DisplayBericht&FIS=200&OBJECT=105070&MODE=METADATA.
Molina, G.A., S.L. Poggio, C.M. Ghersa, 2014: Epigeal arthropod communities in intensively farmed landscapes: effects of land use mosaics, neighbourhood heterogeneity, and field position. Agriculture, Ecosystems & Environment 192, 135-143, DOI: 10.1016/j.agee.2014.04.013.
Monasterolo, M., M.L. Musicante, G.R. Valladares, A. Salvo, 2015: Soybean crops may benefit from forest pollinators. Agriculture, Ecosystems & Environment 202, 217-222. DOI: 10.1016/j.agee.2015.01.012.
Munier-Jolain, N., A. Collard, 2006: Grain legumes and weed management in crop rotations: opportunities and methodologies for reducing environmental impacts of weed control. In: Grain legumes and the environment: how to assess benefits and impacts? AEP (Eds.) Zürich, Switzerland, AEP and FAL, 67-72.
Murphy-Bokern, D., F. Stoddard, C. Watson (Eds.), 2017: Legumes in Cropping Systems. Wallingford, UK, CABI.
Murphy-Bokern, D., J. Dauber, L. Rittler, M. Krön, J. Schuler, M. Reckling, H. Willer, T. Haase, C. Schauman, K. Lindström, C. Zimmer, R. Dewhurst, D. Barkas, J. Recknagel, A. Iancheva, K. Petrovic, S. Alves, T. O’ Donovan, R. Orestis, C. Watson, 2019: Translating knowledge for legume-based farming for food and feed (Legumes Translated). In: First European Conference on Crop Diversification: Book of Abstracts. Messéan, A., D. Drexler, I. Heim, L. Paresys, D. Stilmant, H. Willer (Eds.), Zenodo, 52-53. DOI: 10.5281/zenodo.3516329.
Naeem, S., S. Ahmad, M. Hassan, M. Adil, M.A. Younis, M. Azeem, M.I. Tahir, 2018: Role of pollinators in pea (Pisum sativum) yield at Peshawar valley. Journal of Entomology and Zoology Studies 6, 1280-1282. Access: 18. Juli 2020, URL: http://www.entomoljournal.com/archives/2018/vol6issue2/PartQ/5-6-113-196.pdf.
Nemecek, T., J.S. von Richthofen, G. Dubois, P. Casta, R. Charles, H. Pahl, 2008: Environmental impacts of introducing grain legumes into European crop rotations. European Journal of Agronomy 28, 380-393, DOI: 10.1016/j.eja.2007.11.004.
Newell Price, J.P., D. Harris, M. Taylor, J.R. Williams, S.G. Anthony, D. Duethmann, R.D. Gooday, E. Lord, B.J. Chambers, D.R. Chadwick, T.H. Misselbrook, 2011: An Inventory of Mitigation Methods and Guide to their Effects on Diffuse Water Pollution, Greenhouse Gas Emissions and Ammonia Emissions from Agriculture, Prepared as part of Defra project WQ0106. Access: 18. Juli 2020, URL: https://www.feedadviserregister.org.uk/latest-documents/mitigation-methods--user-guide/mitigationmethods-userguidedecember2011final.pdf.
Noack, M., 2016: Optimierung von Inokulationsmethoden mit Rhizoctonia solani und Sclerotinia sclerotiorum bei Sojabohne und Zuckerrübe. Masterarbeit, Universität für Bodenkultur Wien, 101 S., Access: 18. Juli 2020, URL: https://permalink.obvsg.at/AC13226671.
Nuruzzaman, M., H. Lambers, M.D.A. Bolland, E.I. Veneklaas, 2005: Phosphorus benefits of different legume crops to subsequent wheat grown in different soils of Western Australia. Plant and Soil 271, 175-187, DOI: 10.1007/s11104-004-2386-6.
Olesen, J.E., M.A. Askegaard, I.A. Rasmussen, K. Kristensen, 2011: Crop rotation and crop management effects on cereal yields in arable organic farming in Denmark. In: Organic is Life - Knowledge for Tomorrow. Organic Crop Production. Neuhoff D. (Ed.) Proceedings of the Third Scientific Conference of the International Society of Organic Agriculture Research 1, 280-283.
O'Rourke, M.E., M. Liebman, M.E. Rice, 2008: Ground beetle (Coleoptera: Carabidae) assemblages in conventional and diversified crop rotation systems. Environmental Entomology 37, 121-130, DOI: 10.1603/0046-225X(2008)37[121:GBCCAI]2.0.CO;2.
Osler, G.H., P.C. Van Vliet, C.S. Gauci, L.K. Abbott, 2000: Changes in free living soil nematode and micro-arthropod communities under a canola–wheat–lupin rotation in Western Australia. Soil Research 38, 47-60, DOI: 10.1071/SR99050.
Pahlmann, I., H. Kage, 2018: Steigerung der N-Effizienz im Ackerbau: Welche Optionen haben wir in intensiven Anbausystemen. In: Vorträge zur Hochschultagung 2018 "Landwirtschaft und Ernährung im Spannungsfeld zwischen Umwelt, Gesellschaft und Politik". Schriftenreihe der Agrar- und Ernährungswissenschaftlichen Fakultät der Universität Kiel, 125, 113-125.
Pahlmann, I., K. Sieling, H. Kage, 2018: Anpassung der Fruchtfolge zur Steigerung der N-Effizienz und Reduktion der N-Bilanzsalden im intensiven Ackerbau. Mitteilungen der Gesellschaft für Pflanzenbauwissenschaften 30, 100-101.
Palmer, R.G., P.T. Perez, E. Ortiz-Perez, F. Maalouf, M.J. Suso, 2009: The role of crop-pollinator relationships in breeding for pollinator-friendly legumes: from a breeding perspective. Euphytica 170, 35-52, DOI: 10.1007/s10681-009-9953-0.
Peltonen-Sainio, P., J.K. Niemi, 2012: Protein crop production at the northern margin of farming: to boost or not to boost. Agricultural and Food Science 21, 370-383, DOI: 10.23986/afsci.6334.
Plaza-Bonilla, D., J.M. Nolot, S. Passot, D. Raffaillac, E. Justes, 2016: Grain legume-based rotations managed under conventional tillage need cover crops to mitigate soil organic matter losses. Soil and Tillage Research 156, 33-43, DOI: 10.1016/j.still.2015.09.021.
Poeplau, C., A. Don, 2015: Carbon sequestration in agricultural soils via cultivation of cover crops – A meta-analysis. Agriculture, Ecosystems & Environment 200, 33-41, DOI: 10.1016/j.agee.2014.10.024.
Potts, S., J.C. Biesmeijer, C. Kremen, P. Neumann, O. Schweiger, W.E. Kunin, 2010: Global pollinator declines: trends, impacts and drivers. Trends in Ecology & Evolution 25, 345-353, DOI: 10.1016/j.tree.2010.01.007.
Preissel, S., M. Reckling, N. Schläfke, P. Zander, 2015: Magnitude and farm-economic value of grain legume pre-crop benefits in Europe: A review. Field Crops Research 175, 64-79, DOI: 10.1016/j.fcr.2015.01.012.
Preissel, S., M. Reckling, J. Bachinger, P. Zander, 2017: Introducing legumes into European cropping systems: farm-level economic effects. In: Legumes in cropping systems. Murphy-Bokern, D., F.L. Stoddard, C.A. Watson (Eds.), Wallingford, UK, CABI, 209-225, DOI: 10.1079/9781780644981.0244.
Rees, R.M., J.A. Baddeley, A. Bhogal, B.C. Ball, D.R. Chadwick, M. Macleod, A. Lilly, V. A. Pappa, R.E. Thorman, C.A. Watson, J.R. Williams, 2013: Nitrous oxide mitigation in UK agriculture. Soil Science and Plant Nutrition 59, 3-15, DOI: 10.1080/00380768.2012.733869.
Riedesser, K., 2012: Sommerungen 2012 - Welche Kultur lässt den höchsten Deckungsbeitrag erwarten? Access: 18. Juli 2020, URL: https://www.lwk-rlp.de/de/aktuelles/detail/news/detail/News/sommerungen-2012-welche-kultur-laesst-den-hoechsten-deckungsbeitrag-erwarten/.
Rochester, I.J., M.B. Peoples, N.R. Hulugalle, R.R. Gault, G.A. Constable, 2001: Using legumes to enhance nitrogen fertility and improve soil condition in cotton cropping systems. Field Crops Research 70, 27-41, DOI: 10.1016/S0378-4290(00)00151-9.
Röper, K., F.F. Gröblinghoff, N. Ruoss, J. Berglar, D. Kramps-Alpmann, K. Korte, U. Ernst, J. Brunotte, R. Well, J. Braun, B.C. Schäfer, 2017: Klimaoptimierte Anpassungsstrategien in der Landwirtschaft, Abschlussbericht, 92 S.
Rühl, G, A. Bramm, J.Greef, 2009: Aspekte des Anbaus von Körnerleguminosen. Journal für Kulturpflanzen 61, 312-317.
Schaack, D., C. Rampold, H. Quaing, L. Scheungraber, 2019: AMI Markt Studie – Strukturdaten im ökologischen Landbau in Deutschland 2018. Bonn, Agrarmarkt Informations-Gesellschaft mbH.
Schäfer, B.C., 2013: Mit Fruchtfolgen Witterungsrisiken trotzen! Praxisnah. Access: 18. Juli 2020, URL: http://www.praxisnah.de/index.cfm/article/7750.html.
Schäfer, B.C., N. Lütke-Entrup, 2009: Ökonomische Bewertung von Bodenbearbeitungssystemen in Fruchtfolgen mit Körnerleguminosen. Journal für Kulturpflanzen 61, 309-311, DOI: 10.5073/JfK.2009.09.03.
Schlathölter, M., 2015: Welche Art was mitbringt. DLG Mitteilungen 6, 54-56.
Schlathölter, M., P.H. Petersen, 2015: Wer mit wem. DLG Mitteilungen 3, 56-59.
Schmidt, O., R.O. Clements, G. Donaldson, 2003: Why do cereal–legume intercrops support large earthworm populations? Applied Soil Ecology 22, 181-190, DOI: 10.1016/S0929-1393(02)00131-2.
Schmidt, H., J. Fuchs, D. Möller, D. Wolf, 2014: Körnerleguminosen und Bodenfruchtbarkeit - Kapitel 1: Schlagauswahl. In: Körnerleguminosen und Bodenfruchtbarkeit - Strategien für einen erfolgreichen Anbau. BLE (Eds.), Bonn, Germany, 9-15.
Schmidtke, K., G. Lux, 2015: Wirkung verschiedener Verfahren der Schwefeldüngung auf Ertragsleistung und Vorfruchtwert von Körnerleguminosen im Ökologischen Landbau. BÖLN-Abschlussbericht der Projekte 11OE110 (LWK Niedersachsen) und 11OE111 (HTW Dresden), Access: 18. Juli 2020, URL: https://orgprints.org/29783/.
Schneider, R., E. Heiles, G. Salzeder, K. Wiesinger, M. Schmidt, P. Urbatzka, 2012: Auswirkungen unterschiedlicher Fruchtfolgen im ökologischen Landbau auf den Ertrag und die Produktivität. In: Angewandte Forschung und Beratung für den ökologischen Landbau in Bayern. Ökolandbautag 2012. Wiesinger, K., K. Cais (Eds.). Freising, Germany, Schriftenreihe der LfL, 87-93.
Schulz, K., G. Everwand, J. Dauber, 2019: Faba bean (Vicia faba L.) in wheat-dominated cropping systems: spill-over effects on densities and parasitism rates of vegetation dwelling natural pest control agents? In: First European Conference on Crop Diversification: Book of Abstracts. Messéan, A., D. Drexler, I. Heim, L. Paresys, D. Stilmant, H. Willer (Eds.), Zenodo, 115-116, DOI: 10.5281/zenodo.3516329.
Seidenglanz, M., I. Hunady, J. Poslušná, A.K. Løes, 2011: Influence of intercropping with spring cereals on the occurrence of pea aphids (Acyrthosiphon pisum Harris, 1776) and their natural enemies in field pea (Pisum sativum L.). Plant Protection Science 47, 25-36, DOI: 10.17221/40/2010-PPS.
Shah, A., M. Askegaard, I.A. Rasmussen, E.M.C. Jimenez, J.E. Olesen, 2017: Productivity of organic and conventional arable cropping systems in long-term experiments in Denmark. European Journal of Agronomy 90, 12-22, DOI: 10.1016/j.eja.2017.07.001.
Shuxian, L., P. Chen, 2012: Resistance to Phomopsis Seed Decay in Soybean. Hindawi Publishing Corporation ISRN Agronomy, Article ID 738379, DOI: 10.1155/2013/73 8379.
Sieling, K., O. Christen, 2015: Crop rotation effects on yield of oilseed rape, wheat and barley and residual effects on the subsequent wheat. Archives of Agronomy and Soil Science 61, 1531-1549, DOI: 10.1080/03650340.2015.1017569.
Sillero, J.C., A.M. Villegas-Fernandez, J. Thomas, M.M. Rojas-Molina, A.A. Emeran, M. Fernandez-Aparicio, D. Rubiales, 2010: Faba bean breeding for disease resistance. Field Crops Research 115, 297-307, DOI: 10.1016/j.fcr.2009.09.012.
Smith, R.G., K.L. Gross, S. Januchowski, 2005: Earthworms and weed seed distribution in annual crops. Agriculture, Ecosystems & Environment 108, 363-367, DOI: 10.1016/j.agee.2005.01.014.
Steffens, D., R. Stamm, F. Yan, G. Leithold, S. Schubert, 2005: Rohphosphatmobilisierung von Sommerweizen, Weißlupine und Ackerbohne in einer Fruchtfolge. In: Beiträge zur 8. Wissenschaftstagung zum Ökologischen Landbau. Heß J., G. Rahmann (Eds.), Kassel, Kassel University Press GmbH, 213-216.
Stein, S., H.-H. Steinmann, 2018: Identifying crop rotation practice by the typification of crop sequence patterns for arable farming systems – A case study from Central Europe. European Journal of Agronomy 92, 30-40, DOI: 10.1016/j.eja.2017.09.010.
Steinmann, H.-H., E.S. Dobers, 2013: Spatio-temporal analysis of crop rotations and crop sequence patterns in Northern Germany: potential implications on plant health and crop protection. Journal of Plant Diseases and Protection 120, 85-94, DOI: 10.1007/BF03356458.
Stoddard, F.L., A.H. Nicholas, D. Rubiales, J. Thomas, A.M. Villegas-Fernandez, 2010: Integrated pest management in faba bean. Field Crops Research 115, 308-318, DOI: 10.1016/j.fcr.2009.07.002.
StoffBilV, 2017: Verordnung über den Umgang mit Nährstoffen im Betrieb und betriebliche Stoffstrombilanzen (Stoffstrombilanzverordnung – StoffBilV). Access_ 18. Juli 2020, URL: https://www.gesetze-im-internet.de/stoffbilv/StoffBilV.pdf.
Stute, I., B.C. Schäfer, 2018: Wertvoll für die Fruchtfolge. Raps Spezial Körnerleguminosen 3, 3-5.
Tinivella, T., L.M. Hirata, M.A. Celan, S.A.I. Wright, T. Amein, A. Schmitt, E. Koch, J.M. van der Wolf, S.P.C. Groot, D. Stephan, A. Garibaldi, M.L. Gullino, 2009: Control of seed-borne pathogens on legumes by microbial and other alternative seed treatments. European Journal of Plant Pathology 123, 139-151, DOI: 10.1007/s10658-008-9349-3.
Urbatzka, P., R. Beck, 2015: Entwicklung der Humusgehalte und der Humusqualität in verschiedenen Fruchtfolgen des ökologischen Landbaus. In: 13. Wissenschaftstagung Ökologischer Landbau – Am Mut hängt der Erfolg - Rückblicke und Ausblicke auf die ökologische Landbewirtschaftung. Häring, A.M., et al. (Eds.), Berlin, Germany, Verlag Dr. Köster, 118-122.
Vereijken, P., 1997: A methodical way of prototyping integrated and ecological arable farming systems (I/EAFS) in interaction with pilot farms. European Journal of Agronomy 7, 235-250, DOI: 10.1016/S0378-519X(97)80029-3.
von Richthofen, J.S., H. Pahl, P. Casta, A. Lafarga, T. Nemecek, J.B. Pedesrsen, 2006a: Economic impact of grain legumes in European crop rotations. Grain Legumes 45, 6-19.
von Richthofen, J.S., H. Pahl, D. Bouttet, P. Casta, C. Cartrysse, R. Charles, A. Lafarga, 2006b: What do European farmers think about grain legumes. Grain Legumes 45, 4-15.
Wang, L., H. Cutforth, R. Lal, Q. Chai, C. Zhao, Y.T. Gan, K.H.M. Siddique, 2018: 'Decoupling' land productivity and greenhouse gas footprints: A review. Land Degradation and Development 29, 4348-4361, DOI: 10.1002/ldr.3172.
Wehling, P., 2009: Anbau und Züchtung von Leguminosen in Deutschland - Sachstand und Perspektiven. Journal für Kulturpflanzen 61 (9), 359-364, DOI: 10.5073/JfK.2009.09.11.
Weitbrecht, B., H. Pahl, 2000: Lohnt sich der Anbau von Körnerleguminosen? Ökologie & Landbau 116, 39-41.
Westphal, C., I. Steffan-Dewenter, T. Tscharntke, 2003: Mass flowering crops enhance pollinator densities at a landscape scale. Ecology Letters 6, 961-965, DOI: 10.1046/j.1461-0248.2003.00523.x.
Wilbois, K.P., H. Böhm, B. Bohne, R. Brandhuber, C. Bruns, M. Demmel, M. Finckh, J. Fuchs, A. Gronle, O. Hensel, J. Heß, R. Jörgensen, G.Lux, P. Mäder, D. Möller, H. Schmidt, K. Schmidtke, A.K. Spiegel, L. Tamm, W. Vogt-Kaute, M.Wild, D. Wolf, 2013: Steigerung der Wertschöpfung ökologisch angebauter Marktfrüchte durch Optimierung des Managements der Bodenfruchtbarkeit, BÖLN-Abschlussbericht. Frankfurt am Main, Forschungsinstitut für biologischen Landbau (FiBL), 566 S.
Woodcock, B.A., J. Savage, J.M. Bullock, M. Nowakowski, R. Orr, J.R.B. Tallowin, R.F. Pywell, 2014: Enhancing floral resources for pollinators in productive agricultural grasslands. Biological Conservation 171, 44-51, DOI: 10.1016/j.biocon.2014.01.023.
Xue, A.G., M.J. Morrison, E. Cober, T.R. Anderson, S. Rioux, G.R. Ablett, I. Rajcan, R. Hall, J.X. Zhang, 2007: Frequency of isolation of species of Diaporthe and Phomopsis from soybean plants in Ontario and benefits of seed treatments. Canadian Journal of Plant Pathology 29, 354-364, DOI: 10.1080/07060660709507481.
Young, O.P., G.B. Edwards, 1990: Spiders in United States field crops and their potential effect on crop pests. Journal of Arachnology 18, 1-27.
Zerhusen-Blecher, P., B.C. Schäfer, 2013: Stand des Wissens und Ableitung des Forschungsbedarfes für eine nachhaltige Produktion und Verwertung von Ackerbohne und Erbse. Access: 18. Juli 2020, URL: http://www.orgprints.org/23003/1/23003-12NA118-FHSuedwestfalen-schaefer-2013-forschungsbedarf-ackerbohne-erbse.
Zerhusen-Blecher, P., K. Stevens, B.C. Schäfer, J. Braun, 2018: DemoNet Erbse/Bohne. Access: 18. Juli 2020, URL: http://www.demoneterbo.agrarpraxisforschung.de/index.php?id=301.
Zhong, Z., R.L. Lemke, L.M. Nelson, 2009: Nitrous oxide emissions associated with nitrogen fixation by grain legumes. Soil Biology and Biochemistry 41, 2283-2291, DOI: 10.1016/j.soilbio.2009.08.009.
Zilles, P., 2010: Futtererbsen, eine Anbaualternative zu Braugerste. Dienstleistungszentren Ländlicher Raum Rheinland-Pfalz. Access: 18. Juli 2020, URL: http://www.dlr.rlp.de/Internet/global/themen.nsf/ALL/edee880a3a786e65c125771100415795?OpenDocument&ExpandSection=6.4 %2C6.3 %2C6.2 %2C6.1# Section6.4.
Zimmer, S., T. Haase, H.P. Piepho, E. Stoll, H. Heidt, T. Bohn, J. Heß, 2016: Evaluation of grain legume cropping systems for animal fodder potential and impacts on subsequent wheat yield under less favourable soil conditions in organic agriculture in Luxembourg. Journal of Cultivated Plants 68 (6), 164-174, DOI: 10.5073/JFK.2016.06.02.

 Suchen
Suchen