
Journal für Kulturpflanzen, 74 (11-12). S. 242–256, 2022 | DOI: 10.5073/JfK.2022.11-12.04 | Flachowsky et al.
Der Einfluss der Mendelgenetik auf die Sortenzüchtung bei Apfel und Süßkirsche
The impact of Mendelian genetics on the breeding of apple and sweet cherry
 | Der Autor/Die Autorin 2022 Dies ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Namensnennung 4.0 International Lizenz (CC BY 4.0) zur Verfügung gestellt wird (https://creativecommons.org/licenses/by/4.0/deed.de). |
Zur Veröffentlichung eingereicht/angenommen: 27. Juli 2022/7. November 2022 |
Mendels Einfluss auf die moderne Obstzüchtung ist unverkennbar. Die von ihm aufgestellten Vererbungsregeln haben es Züchtern ermöglicht, Kreuzungsprogramme gezielt zu designen, Vorhersagen über den möglichen Erfolg zu treffen und die Selektion von Nachkommen mit verbesserten Merkmalskombinationen mithilfe molekulargenetischer Marker kostengünstig und effizient zu gestalten. Am Beispiel des Apfels und der Süßkirsche als Vertreter für Kern- und Steinobst werden ausgewählte mendelnde Merkmale vorgestellt, die von besonderer Bedeutung für die Züchtung sind. Darüber hinaus wird der derzeitige Stand an molekularen Markern und genetischen Karten bei diesen Kulturen präsentiert. Sie beeinflussen die Effizienz in der Obstzüchtung enorm. Ihre Entwicklung wäre jedoch ohne Mendels Erkenntnisse nicht möglich gewesen. Beim Apfel, der bedeutendsten einheimischen Obstart, gibt es eine Vielzahl von wirtschaftlich interessanten mendelnden Merkmalen. Einige von ihnen, wie der Säulenwuchs, die rote Fruchtfleischfarbe, das Kältebedürfnis zum Brechen der Winterknospenruhe, die Samenlosigkeit der Früchte sowie gefüllte Blüten für den Anbau als Ziergehölz und die Resistenz gegenüber Schorf und Mehltau sind phänotypisch einfach zu erfassen. Eine Selektion auf der Basis des Phänotyps ist bei diesen Merkmalen meist problemlos möglich, wenngleich sie mithilfe molekularer Marker noch effektiver gestaltet werden kann. Andere Merkmale, wie die Resistenz gegenüber Insekten, verschiedenen Lagerkrankheiten oder der bakteriellen Feuerbrandkrankheit sind nicht so einfach anhand des Phänotyps zu bestimmen. Hier sind für eine erfolgreiche Züchtung molekulare Marker unabdingbar. Das gilt auch für die rote Färbung der Fruchtschale. Bei Süßkirschen ist die Situation sehr ähnlich. Zu den mendelnden Merkmalen mit ökonomischer Bedeutung gehören hier neben der Selbstfertilität auch die Mehltauresistenz und die Farbe der Fruchtschale.
Mendel, Obstzüchtung, Malus domestica, Prunus avium, monogen, Marker
The influence of Mendel on modern fruit breeding is undeniable. The inheritance rules he established have enabled breeders to design breeding programmes in a targeted manner to make predictions about possible success and to select offspring with improved trait combinations using molecular genetic markers in a cost-effective and efficient way. Using the example of the apple and the sweet cherry as representatives of pome and stone fruit, we highlight selected Mendelian traits that are of particular importance for breeding. In addition, the current status of molecular markers and genetic maps and their enormous influence in efficient fruit breeding are presented. However, molecular marker progress would not have been possible without Mendel's insights. In apples, the most important native fruit species, there is a large number of economically interesting Mendelian traits some of which are easy to phenotype. For example, columnar growth, red flesh colour, cold requirement for breaking winter bud dormancy, seedlessness of the fruits and double flowers for cultivation as an ornamental tree and resistance to scab and powdery mildew. Other traits, such as resistance to insects, and the bacterial disease fire blight and various storage diseases are not as straightforward to phenotype. This also applies to red colouration of the fruit skin. Nevertheless, in both situations, molecular markers are indispensable for successful breeding. This also applies to sweet cherries. In addition to self-fertility, powdery mildew resistance and fruit skin colour are among the Mendelian traits with economic significance in sweet cherry breeding.
Mendel, fruit breeding, Malus domestica, Prunus avium, monogenic, marker
Kaum ein anderer Naturwissenschaftler hat die Entwicklungen in der Pflanzenzüchtung in den letzten 100 Jahren so stark beeinflusst wie Gregor Johann Mendel. In Hynčice geboren und später als Priester des Augustinerordens und Abt in Brünn (Tschechische Republik) tätig, war Mendel ein vielseitig interessierter Naturforscher, der mit unterschiedlichsten Pflanzenarten experimentierte. So studierte er beispielsweise bei Obstgehölzen die Ausprägung von Fruchtmerkmalen und stellte dafür umfassende Kreuzungsprogramme auf (Tab. 1). Für die dabei erzeugten Neuzüchtungen wurde er im September 1883 auf der Nationalen Obstausstellung in Brünn vom k. k. Österreichischen Pomologenverein mit einer Medaille der k. k. Wiener Gartenbau-Gesellschaft ausgezeichnet (Orel & Vavra, 1968; Vavra & Orel, 1971).
Tab. 1. Beispiel für ein von Mendel durchgeführtes Kreuzungsprogramm bei Apfel. Die Aufzeichnungen zu diesem Kreuzungsprogramm hat Gregor Mendel handschriftlich in die 1859 erschienene Ausgabe des „Illustrierten Handbuches der Obstkunde“ von F. Jahn, E. Lucas und J.G.G. Oberdiek notiert (Orel & Vavra 1968).
Kombination | Muttersorte | Vatersorte |
1 | Weißer Astrachan | Sommergewürzapfel |
2 | Weißer Astrachan | Der Köstlichste (Bozen) |
3 | Burchardt’s Carolin | Weißer Astrachan |
4 | Alantapfel | Weißer Winterkalvill |
5 | Willy* | Weißer Winterkalvill |
6 | Schneekalvill | Garibaldis Kalvill |
7 | Winterpostoph | Weißer Winterkalvill |
8 | Kanada Renette | Weißer Winterkalvill |
9 | Graue Französische Renette | Süßer Holaart |
10 | Graue Französische Renette | Roter Stettiner |
11 | Graue Französische Renette | Kasseler Renette |
12 | Graue Französische Renette | Der Köstlichste (Bozen) |
13 | Graue Französische Renette | Weißer Winterkalvill |
14 | Graue Französische Renette | Edelborsdorfer |
15 | Graue Französische Renette | Roter Wintertaubenapfel |
16 | Graue Französische Renette | Alantapfel |
17 | Süßer Holaart | Champagner Renette |
18 | Süßer Holaart | Kasseler Renette |
19 | Süßer Holaart | Roter Stettiner |
20 | Süßer Holaart | Weißer Wintertaffetapfel |
21 | Süßer Holaart | Edelborsdorfer |
22 | Süßer Holaart | Weißer Winterkalvill |
23 | Süßer Holaart | Roter Osterkalvill |
24 | Champagner Renette | Sommergewürzapfel |
25 | Champagner Renette | Süßfranke |
26 | Dietzer Goldrenette | Orleans Renette |
27 | Muskatrenette | Weißer Astrachan |
28 | Muskatrenette | Oberdiecks Renette |
29 | Muskatrenette | Edelborsdorfer |
30 | Schneekalvill | Weißer Wintertaffetapfel |
* Bei dieser Sorte scheint es sich vermutlich um die Sorte 'Willy’s Renette' zu handeln
Von wesentlich größerer Bedeutung sind jedoch seine 1866 veröffentlichten Ergebnisse aus Kreuzungsversuchen an Erbsen (Mendel, 1866). Diese führten nach ihrer Wiederentdeckung im Jahr 1900 zu einem Wendepunkt in der modernen Wissenschaft der Genetik (Hossfeld et al., 2017). Mithilfe von Mendels Erkenntnissen war es Wissenschaftlern zu dieser Zeit erstmals möglich, die Vererbung von Eigenschaften mathematisch zu beschreiben und die Anzahl der an der Merkmalsausprägung beteiligten Gene zu ermitteln. Die von Mendel entdeckten Regeln, die eine unabhängige Vererbung (freie Rekombination) voraussetzen, wurden in den Jahren nach ihrer Wiederentdeckung in zahlreichen experimentellen Ansätzen überprüft. Die dabei erzielten Ergebnisse führten letztendlich zu unserem heutigen Verständnis über den Aufbau des Genoms eukaryotischer Lebewesen sowie die Rolle von Chromosomen und Genen bei der Vererbung von Merkmalen.
Einer der Wissenschaftler, welcher die Regeln Mendels zu Beginn des 20. Jahrhunderts auf Allgemeingültigkeit prüfte, war Thomas Hunt Morgan. Bei seinen Experimenten an Fruchtfliegen (Drosophila) kam Morgan zu dem Schluss, dass im Gegensatz zu Mendels Annahme nicht alle Merkmale unabhängig voneinander vererbt werden. Morgan konnte zeigen, dass die Augenfarbe von Fliegen von einem Faktor beeinflusst wird, der auf dem X-Chromosom lokalisiert sein muss, welches das weibliche Geschlecht vererbt (Morgan, 1910). Dieses als genetische Kopplung bezeichnete Phänomen beruht auf der Annahme das Gene, die sich in räumlicher Nähe auf einem Chromosom befinden, wesentlich häufiger gemeinsam an Nachkommen vererbt werden als solche, die sich in einem größeren Abstand zueinander oder sogar auf unterschiedlichen Chromosomen befinden. Crossing over-Ereignisse, die zur Trennung (Rekombination) solcher Gene führen, finden wesentlich seltener statt, je kleiner der räumliche Abstand zwischen zwei Genen ist. Diese Erkenntnis ermöglichte in der Folge die räumliche Anordnung von Merkmalen (Genen) auf einzelnen Chromosomen sowie die Erstellung erster genetischer Karten. Die genetische Kartierung erwies sich in den Anfangsjahren jedoch noch als sehr schwierig, da es bei den meisten Organismen an einer ausreichenden Anzahl einfach zu erfassender Merkmale mangelte, die nur von einem oder wenigen Genen bedingt sind. Um diesen Mangel zu beheben, wurden große Anstrengungen unternommen. Das dabei verfolgte Ziel lag vorrangig in der Identifizierung von Hilfsmerkmalen (Markern), die an ein Zielmerkmal gekoppelt, einfach nachzuweisen, im Zuchtmaterial variabel und in ausreichender Anzahl vorhanden sind. Erst mit der Entdeckung der DNA als Träger der Erbinformation (Watson & Crick, 1953; Nirenberg & Matthaei, 1961), den Restriktionsenzymen (Arber & Linn, 1969) sowie der Entwicklung von Methoden zur DNA-Sequenzierung (Maxam & Gilbert, 1977; Sanger et al., 1977) und der Polymerase-Kettenreaktion (Mullis et al., 1986) wurde dieses Problem hinreichend behoben. Vor allem die rasanten Fortschritte bei der Entwicklung molekularbiologischer Methoden zu Beginn des 21. Jahrhunderts haben dazu geführt, dass molekulare Marker und genetische Karten heute zum Standardrepertoire vieler Züchter gehören. Das ist auch bei heimischen Obstarten, wie dem Apfel oder der Süßkirsche, der Fall. Hier wurden in den letzten Jahrzehnten zahlreiche Merkmale genetisch kartiert, die dafür verantwortlichen Gene identifiziert, in Einzelfällen isoliert und mithilfe einer markergestützten Selektion in hochwertiges Zuchtmaterial überführt. Im Folgenden sind einige ausgewählte Merkmale bei Apfel und Süßkirsche beschrieben, die nach den Mendelschen Regeln vererbt werden und zum Teil eine enorme ökonomische Bedeutung im heutigen Obstbau erlangt haben.
Obwohl beim Apfel, der bedeutendsten einheimischen Obstart in Zentraleuropa, die meisten ökonomisch bedeutenden Merkmale (z. B. Ertrag, Fruchtqualität) von einem Zusammenspiel zahlreicher Gene abhängen (oligo- bzw. polygene Eigenschaften), gibt es dennoch einige, die den klassischen Vererbungsregeln Gregor Mendels folgen. Von diesen besitzen einzelne eine enorme ökonomische Bedeutung. Dazu gehören neben zahlreichen Resistenzen gegenüber biotischen Schaderregern (z. B. Pilze, Bakterien, Insekten) auch solche Merkmale, wie die Rotfärbung von Blättern und Fruchtfleisch, der Säulenwuchs und die Samenlosigkeit oder das Kältebedürfnis zum Überwinden der Winterknospenruhe. Nicht alle dieser mendelnden Merkmale sind jedoch phänotypisch leicht zu erfassen. Das hat in der Vergangenheit oft zur fälschlichen Annahme eines polygenen Vererbungsmusters geführt. Ein gutes Beispiel dafür ist die Resistenz gegenüber dem Feuerbrandbakterium. Im Folgenden sind einige ausgewählte Beispiele für mendelnde Merkmale aufgeführt.
Die Zuverlässigkeit der genetischen Kartierung von Merkmalen hängt im Wesentlichen von der Qualität der zur Verfügung stehenden phänotypischen Daten ab. Nicht alle Merkmale sind in jedem Fall so ausgeprägt, dass sie phänotypisch einfach zu erfassen sind. Ihre Ausprägung hängt sowohl von den beteiligten Genen und Allelen, deren Expression und den sie beeinflussenden Umweltbedingungen ab. In vielen Fällen wird die Ausprägung monogener Merkmale noch von weiteren (z. B. unterstützenden) Faktoren beeinflusst. Dieser Einfluss macht es schwierig Spaltungsverhältnisse zweifelsfrei zuzuordnen. Bei anderen Merkmalen wirken selektive Einflüsse, die sich störend auf das Spaltungsverhältnis auswirken. Das trifft z. B. für die Vererbung des Schorfresistenzgens Rvi6 zu und ist auch für die rote Laubfärbung des Apfels beschrieben. So ist die Schorfresistenz Rvi6 z. B. mit sub-letalen Faktoren gekoppelt, welche als Grund für die gestörte Spaltung vermutet werden (Gao & van de Weg, 2006). Die rote Laubfärbung resultiert aus einer Überfunktion der Flavonoidbiosynthese. Von einzelnen Metaboliten dieses Biosyntheseweges (z. B. Flavonole) ist bekannt, dass sie die Fertilität von Pollen beeinflussen und sich auf die Befruchtung und/oder Samenbildung auswirken (Chagne et al., 2007). Im Folgenden werden Beispiele für mendelnde Merkmale erläutert, bei denen die Erfassung des Phänotyps noch verhältnismäßig zuverlässig möglich ist.
Ein monogenes Merkmal mit einem einfach zu erfassenden Phänotyp, bei dem eine phänotypische Vorselektion des Zuchtmaterials bereits einige Wochen nach der Aussaat möglich ist, ist der Säulenwuchs (Abb. 1). Diese als Columnar bezeichnete Wuchsform wurde zuerst bei 'Wijcik' einer Mutante der Sorte 'McIntosh' entdeckt. Verkürzte Internodien, eine reduzierte Verzweigung und die verstärkte Ausbildung von Kurztrieben an Seitenästen sind für solche Typen charakteristisch. Der dominante Genort (Co) für dieses Merkmal wurde auf Kopplungsgruppe 10 des Apfels kartiert. Die Mutation in 'Wijcik' beruht nach heutigem Kenntnisstand auf einer Insertion eines Ty3/Gypsy-Retrotransposons in den Co-Lokus (Wang et al., 2021). Säulenäpfel spielen heute vor allem bei der Saftproduktion sowie im Hobbyanbau eine Rolle. Die Vorteile dieser Wuchsform werden vor allem in einem geringeren Schnitt- und Pflückaufwand und einer höheren Baumdichte je Flächeneinheit gesehen. Besonderes Interesse hat das Züchtungsprogramm der Hochschule Geisenheim Universität dem Säulenwuchs gewidmet. Die Sorten der „Cats“-Serie, die aus diesem Programm stammen, sind heute im Obstbau weit verbreitet.

Abb. 1. Der Säulenapfel 'PiCol1' aus dem Züchtungsprogramm des JKI wird für den Anbau im Hobbybereich empfohlen. Ursache für das Auftreten dieser Säulenwuchsform ist die Insertion eines 8,2 kbp großen Ty3/Gypsy-Retrotransposons in eine nicht-kodierende Region im Bereich 18.8 Mb auf Kopplungsgruppe 10 des Apfels (Otto et al., 2014). Dieses Retrotransposon führt zur Überexpression des Gens MdCo31 in den Achselknospen säulenförmiger Apfelbäume, welches für eine 2-Oxoglutarat-unabhängige Eisen(II)-Oxygenase kodiert (Wolters et al., 2013).
Eine rote Farbe des Fruchtfleisches ist ein interessantes und lukratives Zuchtziel bei verschiedenen Obstarten (z. B. Apfel, Birne, Kiwi, Zitrusarten). Sie wird vielfach mit einem höheren Gehalt an gesundheitsfördernden Inhaltsstoffen in Verbindung gebracht. Für den Apfel sind bislang zwei Genorte bekannt, die rote (Typ1) bzw. rosa (Typ2) Fruchtfleischfarbe induzieren können (Abb. 2A und B). Bei Genotypen vom Typ1 wird die rote Fruchtfleischfarbe induziert, wenn die regulatorische Region des Transkriptionsfaktorgens MdMYB10 eine sechsfache Wiederholung eines 23 bp (R6) Sequenzmotivs enthält (Espley et al., 2009). Dieses R6-Allel ist autoregulatorisch und fördert die Anthocyan-Synthese. Es wird nach den Mendelschen Regeln vererbt und induziert eine rote Blattfärbung, die besonders in der frühen Wachstumsphase sichtbar ist, sowie eine rote Färbung des Kernhauses. Das Fruchtfleisch kann dabei mehr oder weniger rot gefärbt sein (Volz et al., 2014). Die Selektion auf die rote Laubfärbung (Abb. 2C) erlaubt somit bereits am jungen Sämling eine indirekte Selektion auf rotes Fruchtfleisch. Es gibt inzwischen viele rotfleischige Sorten. Die meisten wie z. B. 'Weirouge' sind für die Vermarktung geschmacklich unzureichend. Mit dem roten Fruchtfleisch wird auch eine bittere Geschmacksnote mitvererbt. Bei neueren Sorten wie 'Baya Marisa', 'Rosette', 'RedLove' und 'Kissabel' ist der Geschmack schon besser. Ob und wie sich diese Sorten am Markt durchsetzen, wird die Zukunft zeigen.

Abb. 2. Unterschiedlich Typen für rote Fruchtfleischfarbe beim Apfel und Möglichkeiten der phänotypischen Frühselektion für rotfleischige Äpfel vom Typ 1. A) Der rotfleischige Zuchtklon PiRo1 aus dem Züchtungsprogramm des JKI ist vom Typ1. Bei den Typ1-rotfleischigen Genotypen ist die rote Färbung von Laub und Fruchtfleisch eine Folge der Insertion eines Minisatellitenmotivs in den Promoterbereich des MdMYB10 Gens des Apfels. B) Der rotfleischige Zuchtklon PiRo3 aus dem Züchtungsprogramm des JKI ist vom Typ2. Bei Typ2-rotfleischigen Äpfeln ist die rote Fleischfarbe eine Folge der Überexpression des Gens MdMYB110a. Die Ursache für diese Überexpression ist unbekannt (Hanke et al., 2020). Im Gegensatz zu Typ1 ist bei den Typ2-Genotypen nur das Fruchtfleisch rot. Fruchtschale und Laub können eine andere Färbung aufweisen. C) Aufgrund der Kopplung zwischen rotem Fruchtfleisch und roter Laubfärbung lassen sich rotfleischige Äpfel vom Typ1 bereits anhand der Laubfärbung der Sämlinge selektieren.
Viele Pflanzen der gemäßigten Klimazonen haben sich an tiefe Wintertemperaturen angepasst. Um diese schadlos zu überstehen, treten sie in eine Phase der Winterruhe (Dormanz) ein, die sich in die drei Teilphasen Para-, Endo- und Ökodormanz untergliedern lässt. Bei der Endodormanz handelt es sich um die tiefe (echte) Winterruhe. In dieser Phase ist das vegetative Wachstum fast vollständig eingestellt und die Knospe auch bei günstigsten Wachstumsbedingungen nicht in der Lage auszutreiben. Um die Endodormanz zu beenden, muss eine sortenspezifische Summe an Kältestunden auf die Knospe wirken. Erst danach tritt sie in die Ökodormanz über. Hier ruht die Knospe noch bis eine sortentypische Summe an Wärmestunden erreicht ist und beginnt dann mit dem Knospenaufbruch.
Äpfel werden unter sehr verschiedenen klimatischen Bedingungen angebaut. Dafür werden angepasste Sorten benötigt. Erfolgt der Anbau in Regionen mit besonders milden Wintern (z. B. Israel), sind Sorten wie 'Anna' geeignet, die ein geringes Kältebedürfnis (~ 300 Kältestunden) haben (Hauagge & Cummins, 1991). Im gemäßigten Klima (z. B. Deutschland) blüht 'Anna' durch das geringere Kältebedürfnis jedoch viel früher als andere Sorten (Abb. 3). Das ist nachteilig, da die zeitige Entwicklung im Frühjahr regelmäßig zu Blütenfrostschäden durch Spätfröste führt. Kürzlich konnte gezeigt werden, dass das Kältebedürfnis des Apfels von einem dominanten Lokus vererbt wird (Hauagge & Cummins, 1991). An diesem Lokus gibt es Gene, die zur Familie der Dormanz-assoziierten MADS-box (DAM)-Gene gehören. Für das Kältebedürfnis scheint vor allem das Gen MdDAM1 von Bedeutung zu sein (Moser et al., 2020; Lempe et al., 2022). In Sorten mit geringem Kältebedürfnis ist MdDAM1 viel schwächer exprimiert als beispielsweise in der Sorte 'Golden Delicious', die ein mittleres Kältestundenbedürfnis besitzt (Moser et al., 2020).

Abb. 3. Die Sorte 'Anna' (A) hat ein geringes Kältebedürfnis und ist in ihrer Blüten- und Blattentwicklung im Frühjahr deutlich früher als beispielsweise die Sorte 'Pinova' (B). Beide Aufnahmen sind am 19. April 2022 auf den Versuchsflächen des Instituts für Züchtungsforschung an Obst in Dresden-Pillnitz entstanden.
Die Blüten der Rosengewächse sind fünfzählig und so hat auch die Apfelblüte im Normalfall fünf Blütenblätter. Es gibt allerdings auch Genotypen, deren Blüten gefüllt sind und deren Anzahl an Blütenblättern ein Vielfaches von fünf ergibt (Abb. 4). Solche Genotypen sind als Zierpflanzen beliebt. Das liegt an ihrem attraktiven Aussehen und daran, dass ihre Blüten meist keine Früchte ausbilden, die im urbanen Raum als störend empfunden werden. Auch bei anderen Rosengewächsen, wie dem Pfirsich gibt es solche Blüten. Hier gibt es zwei Genorte, die diesen Typ bedingen, ein rezessiver Lokus auf Chromosom 2 (Dirlewanger et al., 2004) und ein dominanter Lokus auf Chromosom 6 (Pascal et al., 2017). Solche Arbeiten sind bei Apfel bisher nicht bekannt. Die Entwicklung der Blütenorgane kann mit dem ABC-Modell erklärt werden (Abb. 4E). Um eine Vervielfältigung in der Anzahl an Blütenblättern zu erreichen, gibt es verschiedene Möglichkeiten. Wird zum Beispiel die Aktivität der B-Gene erhöht, vergrößert sich der Bereich, in dem Blütenblätter entstehen und es können mehrere Blütenblattkreise ausgebildet werden. Das ist zum Beispiel beim Pfirsich 'Beijing 2-7' der Fall (Cai et al., 2021). Eine andere Möglichkeit besteht in der Verringerung der Expression eines C-Gens. Dass eine reduzierte Genaktivität der C-Gene AGAMOUS (MdMADS15 und MdMADS22) bei Apfel zu gefüllten Blüten führen kann, wurde bereits gezeigt (Klocko et al., 2016). B- und C-Gene oder solche, die deren Aktivität verändern, sind gute Kandidatengene für die Ausbildung gefüllter Blüten in Apfel.

Abb. 4. Apfelblüten mit mehrfachem Perianth. A) Blüte von Malus pumila MAL003. B) Blüte von Malus spectabilis MAL0544. C) Blüte von Malus hybrida van Eseltine MAL0720. D) Normale Apfelblüte mit einfachem Perianth mit fünf Blütenblättern von Malus domestica 'Pinova'. E) Das ABC-Modell der Blütenbildung. Die Entwicklung der Blütenorgane kann mit dem ABC-Modell erklärt werden. Die zeitlich und örtlich beschränkte Aktivität von Genen mit unterschiedlichen Funktionen (A-, B- und C-Funktion) bestimmt, welche Blütenorgane entstehen. Sind nur Gene mit A-Funktion aktiv, entstehen Kelchblätter. Sind gleichzeitig Gene mit A- und B-Funktion aktiv, entstehen Blütenblätter. Sind Gene mit B- und C-Funktion aktiv, entstehen Staubblätter und Fruchtblätter entstehen, wenn nur Gene mit C-Funktion aktiv sind (Irish, 2017; Coen & Meyerowitz, 1991; Weigel & Meyerowitz, 1994). Gene mit A- und C-Funktion beeinflussen sich aber auch gegenseitig, A-Gene hemmen die Aktivität der C-Gene und umgekehrt.
Die Samenlosigkeit (Parthenokarpie) ist ein Zuchtziel bei vielen Obstarten. Bei Apfel gibt es Sorten, die vorwiegend parthenokarpe Früchte produzieren. Diese Sorten haben Blüten, die weder Blüten- noch Staubblätter besitzen. Anstatt dessen besitzen sie oft zwei Kreise Kelchblätter und zwei bis drei Kreise Fruchtblätter (Abb. 5), (Tobutt, 1994). Durch das Fehlen der Blütenblätter werden keine Insekten zur Bestäubung angelockt. Interessanterweise werden trotz fehlender Befruchtung Früchte ausgebildet. Diese besitzen keine Samen (Abb. 5). Blütenblattlose Apfelblüten wurden schon Mitte des 19. Jahrhunderts beschrieben (Dochnahl, 1855; Ritter Beck von Mannagetta & Abel, 1894). Sorten, die solche Blüten besitzen sind M. pumila 'apetala', M. domestica 'Spencer Seedless', 'Wellington Bloomless' und 'Rae Ime'. In Kreuzungsexperimenten konnte gezeigt werden, dass das Merkmal monogen rezessiv vererbt wird (Tobutt, 1994). Bei dem Gen, welches dieses Merkmal bedingt, handelt es sich mit hoher Wahrscheinlichkeit um das PISTILLATA (MdPI) Gen des Apfels. Dieses Blütenorganidentitätsgen ist weder in 'Spencer Seedless' noch in 'Wellington Bloomless' oder 'Rae Ime' aktiv. In diesen Sorten ist es durch die Insertion eines Transposons im 4. ('Rae Ime') bzw. 6. Intron ('Spencer Seedless' und 'Wellington Bloomless') zerstört worden (Yao et al., 2001). In funktionellen Studien konnte das Fehlen von MdPI als mögliche Ursache der Parthenokarpie bestätigt werden (Tanaka et al., 2007).
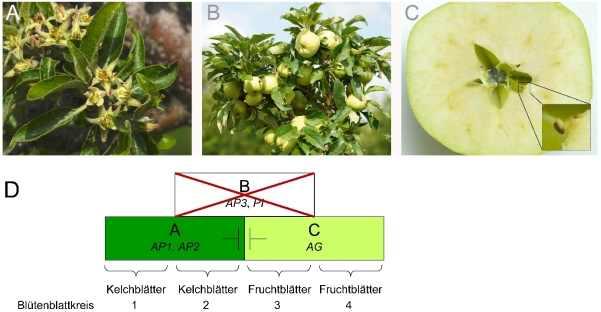
Abb. 5. Samenlosigkeit bei Apfel. A) Blüten von M. pumila 'apetala' bilden keine Blütenblätter aus. B) Dadurch sind sie nicht attraktiv für Bienen und werden nicht bestäubt. Trotz fehlender Befruchtung werden Früchte ausgebildet, sogenannte parthenokarpe Früchte. C) Diese haben jedoch keine Samen. D) Modell einer Blüte mit einer Mutation in einem B-Funktionsgen, die zum Verlust der Genfunktion führt. In den Sorten 'Spencer Seedless', 'Wellington Bloomless' und 'Rae Ime' ist das B-Funktionsgen PISTILLATA des Apfels (MdPI) durch Insertion eines Retrotransposons zerstört worden. Damit sind in den Blütenblattkreisen 2 und 3 ausschließlich A- bzw. C-Funktionsgene aktiv. Folglich werden in diesen Blütenblattkreisen ebenfalls Kelch- bzw. Fruchtblätter gebildet.
Von großer ökonomischer Bedeutung ist die Resistenz gegenüber Apfelschorf. Sie ist schon seit Beginn der wissenschaftlich basierten Obstzüchtung in den 1920er-Jahren ein bedeutendes Zuchtziel. In der Vergangenheit wurde zur Verbesserung der Resistenz vielfach das Resistenzgen Rvi6 aus dem Wildapfel Malus floribunda (Mf821) benutzt. Die von Rvi6 vermittelte Schorfresistenz ist heute in mehr als 100 neu gezüchteten Apfelsorten zu finden. Die Introgression von Rvi6 in den Kulturapfel geht auf eine Kreuzung eines Klons Mf821 mit der Sorte 'Morgenduft' aus den 1910er Jahren zurück (Crandall, 1926). Später in den 1940er-Jahren entdeckte Hough, dass eine Population aus einer Kreuzung von zwei F‘1-Nachkommen dieser Kreuzung nahezu 1:1 für Resistenz und Anfälligkeit spaltete. Weitere Kreuzungen bestätigten die Annahme, dass die Resistenz auf einem dominanten Gen beruht. Dieses Gen wurde zuerst Vf (Venturia floribunda) und später Rvi6 (Resistenz to Venturia inaequalis) genannt (Hough et al., 1953; Bus et al., 2011). Aus der Kreuzung von Hough wurden zwei schorfresistente F´2-Nachkommen ausgewählt und an Züchter weltweit abgegeben. 1975 erhielt die Apfelsorte 'Prima', als erste schorfresistente Apfelsorte mit Vf, Sortenschutz. Auch in Deutschland (Müncheberg) begann Erwin Baur in den 1920er-Jahren mit der Schorfresistenzzüchtung. Als Resistenzdonor nutzte er die Sorte 'Antonovka', die jedoch eine komplexere Vererbung der Schorfresistenz aufweist. Mittlerweile sind ca. 20 Schorfresistenzgene bekannt (Tab. 2), die in der Züchtung genutzt werden können. Für die Resistenzgene wurde ein Rassetestersortiment von Hosts (h) entwickelt, welches in vielen Ländern der Welt angebaut wird, um die Wirksamkeit der Resistenz in unterschiedlichen Umwelten zu testen. Einige der Resistenzen sind monogen, andere beruhen auf zwei oder mehreren Genen. So beruht z. B. die Schorfresistenz von Hansens baccata#2 auf zwei Genen. Die Resistenz von 'Geneva' scheint von vier Genen bedingt zu sein, die des 'Dülmener Rosenapfel' von einem Majorgen und drei Resistenz-QTL, und die von Mf821 von zwei rassespezifischen Resistenzgenen. Einige dieser Schorfresistenzen sind bereits durchbrochen. Deshalb versuchen Züchter heute durch die Kombination (Pyramidisierung) mehrerer Resistenzen eine dauerhafte Widerstandsfähigkeit zu erreichen.
Tab. 2. Aufgeführt sind 20 dominante Schorfresistenzgene mit alter und neuer Bezeichnung und die entsprechenden Hosts (h) im Schorfrassetestersortiment.
Host | Genotyp | Name Resistenzgen | |
neu | alt | ||
h0 | Gala |
|
|
h1 | Golden Delicious | Rvi1 | Vg |
h2 | TSR34T15 | Rvi2 | Vh2 |
h3 | Geneva | Rvi3 | Vh3 |
h4 | TSR33T239 | Rvi4 | Vh4,Vx,Vr1 |
h5 | 9-AR2T196 | Rvi5 | Vm |
h6 | Priscilla | Rvi6 | Vf |
h7 | Malus floribunda 821* | Rvi7 | Vfh |
h8 | B45 | Rvi8 | Vh8 |
h9 | J34 | Rvi9 | Vdg |
h10 | A723-6 | Rvi10 | Va |
h11 | A722-7 | Rvi11 | Vbj |
h12 | Hansen’s Baccata #2 | Rvi12 | Vb |
h13 | Durello di Forli | Rvi13 | Vd |
h14 | Dülmener Rosenapfel | Rvi14 | Vdr1 |
h15 | GMAL 2473 | Rvi15 | Vr2 |
h16 | MIS op.93.051 G07-098b | Rvi16 | Vmis |
h17 | Antonovka APF22 | Rvi17 | Va1 |
h18 | 1980-015-025 | Rvi18 | V25 |
| Honeycrisp | Vhc1** |
|
| Honeycrisp | Vhc15** |
|
* ersetzt bislang eine F1 von Mf821, die nur Rvi7 enthält
** vorläufige Bezeichnung der Resistenzgene
Im Gegensatz zu anderen mendelnden Merkmalen, ist die Resistenz gegenüber dem bakteriellen Feuerbrand phänotypisch nur schwer zu erfassen. Das Spektrum an Symptomen reicht in spaltenden Populationen oft von vollständig resistent (keine sichtbaren Symptome) bis hin zu vollständiger Anfälligkeit. Diese kann im Extremfall zum Absterben der Pflanze führen.
Die Feuerbrandkrankheit wird durch das Bakterium Erwinia amylovora verursacht und ist die bedeutendste Bakterienerkrankung des Kernobstes (Peil et al., 2020). Ihr Erreger gehört in der EU zu den regulierten Nicht-Quarantäneschaderregern (Picard et al., 2018). Für nachweislich infizierte Pflanzen wird meist eine Rodung empfohlen. Künstliche Tests auf Resistenz erfolgen deshalb durch Inokulation frisch veredelter Pflanzen, die ausschließlich zum Zweck der Testung hergestellt und anschließend entsorgt werden. Diese Pflanzen sind juvenil und bilden in den ersten Jahren nach der Veredelung noch keine Blüten (Hanke et al., 2007). Für die künstliche Inokulation werden deshalb meistens junge Triebe genutzt (Peil et al., 2014), obwohl die natürliche Infektion über die Blüten erfolgt. Das Ausmaß der Infektion (Anfälligkeit) wird dann anhand des Verhältnisses zwischen der Gesamtlänge des Triebes und der Länge der sich bildenden Läsion als „Prozentuale Läsionslänge“ (PLL) bestimmt. Dabei steht ein Wert von PLL = 0 für vollständige Resistenz. Pflanzen mit einem Wert von PLL = 10 gelten ebenfalls als resistent, während Werte von PLL > 15 auf Anfälligkeit schließen lassen (Peil et al., 2007; Emeriewen et al., 2021). Für Individuen, die sich phänotypisch an den beiden extremen Enden der Population befinden, ist die Zuordnung einfach. Diese Pflanzen besitzen bei der genetischen Kartierung meist die Markerallele, die mit Resistenz bzw. Anfälligkeit gekoppelt sind, zu beiden Seiten des Resistenzlokus. Für die genaue Bestimmung der Position des Resistenzgens sind jedoch solche Pflanzen entscheidend, bei denen es innerhalb des Lokus zu einer Rekombination gekommen ist (Emeriewen et al., 2018). Diese Pflanzen besitzen auf der einen Seite des Lokus Markerallele, die mit Resistenz gekoppelt sind. Auf der anderen Seite besitzen sie Allele, die mit Anfälligkeit assoziiert sind. Phänotypisch können diese Pflanzen resistent oder anfällig sein (Emeriewen et al., 2018; Fahrentrapp et al., 2013). Für die Zuordnung der entsprechenden Genotypen zum Phänotyp resistent oder anfällig ist der Median einer Population ein geeignetes Kriterium, aber auch nicht immer sicher. Eine genaue Bestimmung des Genortes für das Resistenzgen ist dadurch sehr schwierig. Trotz dieser Schwierigkeiten wurden Kandidatengene für Feuerbrandresistenz in Wildapfelgenotypen, wie M. ×robusta 5 (Fahrentrapp et al., 2013), M. fusca MAL0045 (Emeriewen et al., 2018), M. ×arnoldiana (Emeriewen et al., 2017; Emeriewen et al., 2021) und der Zierapfelsorte 'Evereste' (Parravicini et al., 2011) identifiziert. Diese befinden sich auf den Chromosomen 3, 10 und 12. Molekulare Marker, die mit diesen Resistenzen gekoppelt sind, werden heute für die markergestützte Selektion (MAS) in verschiedenen Züchtungsprogrammen weltweit verwendet.
Neben der Resistenz gegenüber Pilzen und Bakterien ist auch die Widerstandsfähigkeit gegenüber Insekten von großer ökonomischer Bedeutung. Vor allem Apfelblutläuse (Eriosoma lanigerum) zählen neben der Mehligen Apfelblattlaus (Dysaphis plantaginea) und der Rosigen Apfelfaltenlaus (Dysaphis devecta) zu den bedeutendsten Schadinsekten im Kernobstanbau. Die Bekämpfung von Blutläusen ist besonders schwierig, da diese im oberen Wurzelbereich und in Rindenritzen überdauern. Deshalb wurde schon sehr frühzeitig in East Malling (England) mit der Züchtung resistenter Apfelunterlagen begonnen (Crane et al., 1936). In Deutschland lag das Ziel in der Züchtung resistenter Edelsorten. Hier wurden bereits 1919 erste Züchtungsarbeiten in der neu gegründeten Biologischen Reichsanstalt für Land- und Forstwirtschaft in Naumburg/Saale unter der Leitung von Carl Börner initiiert (Jancke, 1937). Im Rahmen seiner Arbeiten konnte Bramstedt (1938) drei Reaktionstypen nach Blutlausbefall identifizieren. Pflanzen mit Blutlausbefall und anschließender Bildung von Gallengewebe wurden als anfällig eingestuft. Befallsfreie Pflanzen wurden als immun/resistent eingestuft. An diesen konnte Bramstedt (1938) als erster histologisch die Bildung von nekrotischem Gewebe an Trieben und Wurzeln nachweisen. Zusätzlich beobachtete er eine intermediäre Gruppe, die im Freiland nur sehr selten auftrat. Pflanzen dieser Gruppe wurden aufgrund der Bildung von nekrotischem Gewebe ebenfalls als resistent eingestuft. Mithilfe dieser Klassifizierung war es möglich geworden, die genetischen Mechanismen der Resistenz in spaltenden Populationen mit resistenten Sorten (z. B. 'Northern Spy', 'Ontario', 'Harberts Renette') und Apfelwildartenakzessionen von M. baccata, M. micomalus, M. coronaria, M. orthocarpa und M. pumila) aufzuklären (Börner & Gollmick, 1943). Gleichzeitig wurden umfangreiche Evaluierungsarbeiten in vorhandenen Sortimenten durchgeführt (Speyer, 1924; Jancke, 1937; Shay, 1962). Bei den Apfelsorten erwies sich vor allem die Resistenz von 'Northern Spy' als besonders stabil. Diese Resistenz wird von dem dominanten Resistenzgen Er1 hervorgerufen (Knight et al., 1962). Später wurden mit Er2 und Er3 weitere monogene Blutlausresistenzen in M. robusta (King et al., 1991; Alston et al., 2000; Bus et al., 2008) und M. sieboldii 'Aotea 1' (Bus et al., 2002) identifiziert. Die Resistenzgene Er1 und Er3 sind bereits von einzelnen Biotypen der Blutlaus überwunden worden (Giliomee et al., 1968; Rock & Zeiger, 1974; Sen Gupta & Miles, 1975). Ob Er2 auch schon gebrochen ist, ist bislang unbekannt (Cummins & Aldwinckle, 1983).
Für die Mehlige Apfelblattlaus konnte in M. robusta mit Smh ebenfalls eine monogene Resistenz identifiziert werden (Alston & Briggs, 1970) und für die Rosige Apfelfaltenlaus wurden mit Sd1, Sd2 und Sd3 drei monogene Resistenzen in 'Coxs Orange Pippin', 'Northern Spy' und Malus × robusta nachgewiesen, welche jeweils gegen unterschiedliche Biotypen des Erregers wirksam sind (Brady, 1991). Für die MAS wurden bereits Marker für die Gene Er1, Er2 und Er3 sowie Sd1, und Sd2 entwickelt (Bus et al., 2008; Cevik & King, 2002; Gardiner et al., 2001; Gardiner et al., 2007). Diese finden heute Anwendung in verschiedenen Züchtungsprogrammen.
Zu den für die Kirschenzüchtung wichtigen Merkmalen gehören u. a. die Fruchtfarbe, die Fruchtgröße, die Fruchtfleischfestigkeit sowie die Resistenz gegenüber biotischen und abiotischen Stressfaktoren (Pijut, 2020). Obwohl die Züchtungsforschung bei Süßkirschen im Vergleich zum Apfel weniger intensiv entwickelt ist, gibt es auch hier inzwischen eine Vielzahl an genetischen Karten sowie erste molekulare Marker. Gerade die Entwicklung von PCR-basierten Methoden und Verfahren der DNA-Sequenzanalyse haben in den letzten Jahrzehnten zur Identifizierung von zahlreichen Selbstinkompatibilitätsallelen sowie zur gezielten Selektion selbstfertiler Genotypen geführt (Dirlewanger et al., 2009; Pijut, 2020). In der Genomdatenbank der Rosengewächse (https://www.rosaceae.org) sind bislang 51 genetische Karten für die Süßkirsche gelistet. Diese wurden mit unterschiedlichen Markertypen (RFPL, SSR und SNP) erzeugt (Pijut, 2020). Blüten- und Fruchteigenschaften, wie die Fruchtgröße und Selbstinkompatibilität, sind klassische Beispiele für mendelnde Merkmale, die sowohl phänotypisch als auch molekulargenetisch mithilfe segregierender Populationen untersucht wurden (Zhang et al., 2010).
Für die Vererbung der Fruchtgröße wurde ein Lokus auf Kopplungsgruppe 2 der Süßkirsche identifiziert (Zhang et al., 2010; Olmstead et al., 2008). Weitere Loci, die mit Fruchtgröße assoziiert sind, wurden auf anderen Kopplungsgruppen gefunden (Rosyara et al., 2013). Kürzlich wurde jedoch gezeigt, dass die Markerallele, die mit dem Lokus auf Kopplungsgruppe 2 assoziiert sind, den größten Einfluss auf das Merkmal Fruchtgröße haben (Zhang et al., 2010). Mithilfe dieser Marker lassen sich die Sorten nach Haplotypen unterteilen, die mit großen und kleinen Früchten assoziiert sind. Damit können für Züchtungsprogramme gezielt Elternsorten ausgewählt werden, die eine hohe Fruchtgröße an ihre Nachkommen vererben. Gleichzeitig ermöglichen diese Marker auch die Selektion von Nachkommen, die das genetische Potenzial haben, große Früchte auszubilden. Heute finden diese Marker in verschiedenen Züchtungsprogrammen Anwendung.
Das Vorkommen von Selbst- und Kreuzungsinkompatibilität bei Pflanzen hat Biologen schon sehr lange interessiert. Ausgehend von den Erkenntnissen von Correns, East, Lehmann und Sirks sowie den Ergebnissen eigener Studien konnten Crane & Lawrence (1929) erstmals die genetischen und zytologischen Ursachen der gametophytischen Selbstinkompatibilität bei Obst beschreiben. Dabei konnten sie zeigen, dass die Inkompatibilität von genetischen Faktoren bestimmt wird, die den von Mendel beschriebenen Vererbungsregeln für einen monogenen Erbgang folgen.
Ursache für die Selbstinkompatibilität bei Süßkirschen ist ein gametophytisches Selbstinkompatibilitätssystem, welches von einem Selbstinkompatibilitätslokus, dem S-Lokus kontrolliert wird (de Nettancourt, 1977; Tehrani & Brown, 1992). Dabei wird die Inkompatibilitätsreaktion durch zwei eng gekoppelte Gene bestimmt, die sich am S-Lokus befinden. Das eine Gen kodiert für eine griffelspezifische Ribonuklease, die sogenannte S-RNase. Das andere Gen kodiert ein pollenspezifisches F-Box-Protein, das SFB-Protein (Yamane et al., 2003). Von beiden Genen gibt es zahlreiche Allele, die als S1-, S2-, S3-Allele usw. bekannt sind. Da die Süßkirsche diploid ist, besitzt sie jeweils zwei Allele der S-RNase und des SFB-Proteins. Bei der Bestäubung keimen die haploiden Pollen auf der Narbe aus und wachsen in das diploide Griffelgewebe ein. Dabei nehmen sie die beiden im Griffel exprimierten S-RNasen auf. Diese werden normalerweise in der Folge abgebaut. Stimmt jedoch das Allel des SFB-Proteins im Pollen mit dem Allel einer der beiden S-RNasen überein, wird diese S-RNase vor dem Abbau geschützt. Sie bleibt funktional und hemmt das Pollenschlauchwachstum im Griffel. Dadurch wird die Befruchtung der Eizelle im Fruchtknoten verhindert. Eine erfolgreiche Befruchtung der Eizelle kann nur erfolgen, wenn sich das S-Allel im Pollen von den beiden S-Allelen im Griffelgewebe unterscheidet. Für das gametophytische Selbstinkompatibilitätssystem der Süßkirsche sind drei verschiedene Pollen-Griffel-Interaktionen möglich (Abb. 6).
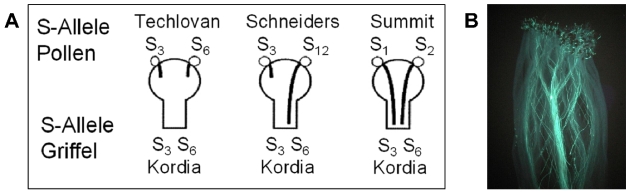
Abb. 6. Möglichkeiten für Pollen-Griffel-Interaktionen bei Süßkirsche, die durch das gametophytische Selbstinkompatibilität bedingt werden. A) Bei der Kombination 'Kordia' × 'Techlovan' kann der Pollen von 'Techlovan' mit den Allelen S3 und S6 die Sorte 'Kordia' (ebenfalls S3 und S6) nicht befruchten, da beide Sorten die gleichen S-Allele besitzen und somit inkompatibel sind. Bei der Kombination 'Kordia' × 'Schneiders' unterscheidet sich das S12-Allel in den Pollen von 'Schneiders' von den beiden S-Allelen im Griffel von 'Kordia'. Im Pollenschlauch des S12-Alleles werden beide S-RNAsen von 'Kordia' abgebaut. Damit kann der Pollenschlauch weiterwachsen und zu einer erfolgreichen Befruchtung führen. In den Nachkommen dieser Kreuzung sind die beiden S-Genotypen S3S12 und S6S12 zu erwarten. In der Kombination 'Kordia' × 'Summit' unterscheiden sich beide S-Allele der Vatersorte (S1S2) von denen der Muttersorte (S3S6). Beide Pollen-S-Genotypen von 'Summit' können 'Kordia' erfolgreich befruchten. In den Nachkommen sind die vier S-Genotypen S1S3, S2S3, S1S6 und S2S6 zu erwarten. B) Narbe mit Pollen der Sorte 'Pieraszt'. Die Situation entspricht dem Fall 'Kordia' × 'Schneiders'. Einige Pollenschläuche können tief in den Griffel einwachsen, während andere bereits nach kurzer Zeit gestoppt werden.
Zu Beginn des 20. Jahrhunderts erfolgte die Bestimmung der Bestäubungseignung mithilfe umfangreicher Bestäubungsversuche (Kobel, 1927; Kobel et al., 1938; Crane & Lawrence, 1929; Crane & Brown, 1937; Brown, 1955; de Vries, 1968; Branscheidt, 1931; Vahl, 1965; Mihatsch & Schumann, 1971; Schmidt et al., 1999). Im Ergebnis dieser Untersuchungen wurden bereits erste Inkompatibilitätsgruppen beschrieben. Eine genaue Identifizierung dieser Gruppen war jedoch nicht möglich, da hauptsächlich mit lokalen bzw. regionalen Sortensortimenten gearbeitet wurde. Zusätzlich erschwerten Synonyme und Falschbezeichnungen einzelner Sorten die exakte Zuordnung. Auf der Grundlage der im John Innes Centre in England begründeten Nomenklatur für die Inkompatibilitätsgruppen bei Süßkirsche beschrieb Knight (1969) bereits 15 Inkompatibilitätsgruppen bei 176 untersuchten Süßkirschen. In der Mitte der 1990er-Jahre identifizierten Bošković & Tobutt (1996) mithilfe von Isoenzymanalysen S-RNase-Allele im Griffelgewebe. Mit dieser Methode war es erstmals möglich, die S-Allele von Süßkirschen im Labor unabhängig vom Baum zu bestimmen. Im Ergebnis dieser Isoenzymanalysen wurden die S-Allel-spezifischen Bandenmuster für die Allele S1–S11 sowie S12-S14 beschrieben (Bošković & Tobutt, 1996; 2001; Bošković et al., 1997).
Mit der fortschreitenden Entwicklung der Molekularbiologie wurde die PCR-Technik für die Bestimmung der S-Allele eingesetzt. Voraussetzung dafür war die Aufklärung der Struktur des S-RNase-Gens (Ushijima et al., 1998). Die kodierende DNA-Sequenz dieses Gens wird von zwei Introns unterbrochen, deren Größe bei den einzelnen S-Allelen variiert. Basierend auf dieser Erkenntnis wurden consensus- und S-Allel-spezifische Primerpaare entwickelt, deren Amplifikationsprodukte diese Introns einschließen (Tao et al., 1999; Wiersma et al., 2001; Sonneveld et al., 2001; 2003; 2006). Diese Entwicklung führte zu einem gewaltigen Fortschritt bei der Identifizierung der S-Genotypen. Eine zusammenfassende Übersicht über Sorten und deren S-Allele ist bei Schuster (2020) zu finden.
Die Kenntnisse über die S-Genotypen einzelner Sorten ermöglichen dem Obstbau die gezielte Auswahl von Süßkirschsorten, die sich gegenseitig befruchten können und eine hohe Ertragsleistung von Neuanlagen gewährleisten. Große Bedeutung haben diese Informationen auch für die Züchtung. Hier kann die Fertilität von Kreuzungskombinationen bereits im Vorfeld bestimmt werden.
Verbunden mit dem Wunsch, die Selbstinkompatibilität von Süßkirschen zu überwinden, wurden ab dem Jahr 1946 am John Innes Centre in Norwich (England) umfangreiche Untersuchungen durchgeführt. Im Ergebnis konnten drei selbstfertile Süßkirschensämlinge selektiert werden (Lewis & Crown, 1954). Zwei dieser Sämlinge (JI 2420 und JI 2434) entstanden durch Bestrahlung des Pollens der Vatersorte mit Röntgenstrahlung. Ein dritter Sämling (JI 2538) ist auf eine spontane Mutation zurückzuführen (Lewis & Crowe, 1954). Mit dem Klon JI 2420 bei dem die Pollenfunktion des S4-Allels (S4') mutiert ist, wurde von Lapins 1956 an der Agriculture Canada Research Station in Summerland (Kanada) ein Zuchtprogramm zur Erzeugung selbstfertiler Süßkirschen begonnen. Im Rahmen dieses Programms wurde 1970 mit 'Stella' die weltweit erste selbstfertile Sorte selektiert (Lapins, 1971). Diese Sorte wurde fortan in vielen Züchtungsprogrammen verwendet. Die beiden 1971 von Lane und Schmid in Summerland (Kanada) selektierten Süßkirschensorten 'Lapins' und 'Sunburst' waren die ersten selbstfertilen Sorten die im Erwerbsobstbau genutzt wurden (Lane & Schmid, 1984). Weltweit wurden bis zum Jahr 2020 91 selbstfertile Süßkirschensorten beschrieben (Schuster, 2020). Im Zuchtprozess lassen sich durch die monogene Vererbung der S4'-Pollenmutation Kreuzungspopulationen mit 100 % bzw. 50 % selbstfertilen Nachkommen planen (Tab. 3).
Tab. 3. Prozentualer Anteil an selbstfertilen Nachkommen (SF) bei Kreuzungen mit selbstfertilen Elternsorten mit dem mutiertem S4-Allel S4'. Bei Kreuzungskombinationen mit einem selbstfertilen Elternpartner erhält man 100 % selbstfertile Nachkommen, wenn beide Elternsorten ein gleiches S-Allel besitzen. Alle Nachkommen einer solchen Kreuzung haben den gleichen S-Genotyp. Unterscheiden sich die S-Allele von Mutter- und Vatersorte, so ist mit 50 % selbstfertilen Nachkommen zu rechnen. Die Nachkommen spalten in vier unterschiedlich S-Genotypen auf.
S-Genotyp | S-Genotyp | S-Genotyp | SF Nachkommen |
S3S12 | S3S4' | S4'S12 | 100 |
S1S4' | S1S2 | S2S4' | 100 |
S1S2 | S3S4' | S1S3/S2S3 | 50 |
S1S4' | S4'S9 | S1S4'/S1S9 | 75 |
Die von Gregor Mendel aufgestellten Regeln zur Vererbung von Merkmalen haben die Obstzüchtung im vergangenen Jahrhundert stark beeinflusst. Bei unterschiedlichen Obstarten konnten Werteigenschaften identifiziert werden, deren Vererbung sich mit Mendels Regeln erklären lässt. Für solche Merkmale ist der Erfolg einer Kreuzung vorhersagbar geworden. Einige dieser Merkmale, wie z. B. Schorfresistenz, Rotfleischigkeit und Säulenwuchs bei Apfel, sowie Fruchtgröße und Selbstfertilität bei Süßkirschen, haben ein großes Marktpotenzial erlangt. Neue Sorten, die aus gezielten Kreuzungsprogrammen unter Anwendung der Mendelschen Vererbungsregeln entstanden sind, erobern zunehmend den Markt und verdrängen nach und nach Topsorten wie 'Golden Delicious' und 'Jonagold' (Schröpfer et al., 2022), die einst als Zufallssämlinge entstanden sind, oder aus den frühen Anfängen der Kreuzungszüchtung stammen. Allein mit der Schorfresistenz Rvi6 aus Malus floribunda 821 sind weltweit mehr als 100 neue Apfelsorten entstanden. Von diesen hat vor allem die Sorte 'Topaz' den Sprung unter die Topsorten in Europa geschafft. Andere Rvi6-resistente Apfelsorten, wie 'Rustica', 'Natyra', 'Ariane' und 'Bonita' werden derzeit für den Anbau in der biologischen Produktion empfohlen (Friedli et al., 2021). Zunehmend entstehen in den Züchtungsprogrammen aber auch schorfresistente Apfelsorten, deren Resistenz auf einem anderen Resistenzmechanismus beruht (Flachowsky & Töpfer, 2021). Ein gutes Beispiel dafür ist die Sorte 'Pia 41' aus der Züchtung des JKI. Diese Sorte, deren Resistenz aus der Sorte 'Honey Crisp' stammt, findet derzeit einen wachsenden Zuspruch in der Direktvermarktung. Sorten mit Kombinationen von zwei und mehr Resistenzgenen, wie 'Rea Bellina' (Rvi2 und Rvi4) stoßen bereits jetzt auf Interesse in einigen Ländern Osteuropas. Auch bei Süßkirschen ist ein solcher Sortenwechsel zu verzeichnen. Hier ist das Interesse an neuen großfrüchtigen Sorten mit festem Fruchtfleisch und hervorragendem Geschmack in den letzten Jahren stark gestiegen. Sorten wie 'Areko' stehen heute in vielen Ländern weltweit in der Testung bzw. schon im Anbau. Andere Zuchtklone bahnen sich aufgrund ihrer Selbstfertilität und der hervorragenden Fruchteigenschaften ihren Weg (Abb. 7).

Abb. 7. Neue Sorten aus dem Züchtungsprogramm des JKI. A) Die neue Apfelsorte 'Pia 41' aus dem Züchtungsprogramm des JKI trägt eine Schorfresistenz aus der Sorte 'Honey Crisp'. B) Die Sorte 'Rea Bellina' des JKI besitzt eine Kombination der beiden Schorfresistenzen Rvi2 und Rvi4. C) Die Süßkirschensorte 'Areko' des JKI zeichnet sich aus durch eine überdurchschnittliche Fruchtgröße, einen hervorragenden Geschmack und eine hohe Fruchtfestigkeit. D) Selbstfertiler Zuchtklon bei Süßkirsche mit vielversprechenden Frucht- und Lagereigenschaften.
Neben der Wiederentdeckung der Mendelschen Vererbungsregeln hat die Züchtung von Kern- und Steinobst auch von vielen anderen Entwicklungen in der Genetik profitiert. Vor allem durch die neuen Methoden im Bereich der Genomanalyse hat der Erkenntnisgewinn noch einmal drastisch zugenommen. So wurden in den vergangenen drei Jahrzehnten zahlreiche genetische Karten für biparentale Populationen erstellt, die für unterschiedlichste Merkmale segregieren. Die dabei identifizierten Marker, die eng an Werteigenschaften gekoppelt sind, werden heute in Züchtungsprogrammen zur markergestützten Selektion genutzt (Hanke et al., 2020). Ein Ziel der markergestützten Selektion bei Apfel ist die Pyramidisierung von Resistenzgenen gegenüber Apfelschorf, Mehltau, Feuerbrand und verschiedenen Schadinsekten (Flachowsky & Töpfer, 2021). Am Institut für Züchtungsforschung an Obst des JKI in Dresden-Pillnitz werden dazu Marker für acht Schorf-, vier Mehltau- und vier Feuerbrandresistenzen genutzt, um unterschiedliche Pyramiden und Kombinationen von Resistenzgenen anhand des Genotyps zu selektieren. Eine solche zielgerichtete Kombination von Resistenzen, die gegen den gleichen Erreger gerichtet sind, ist alleine auf der Grundlage phänotypischer Verfahren nicht zu leisten.
Für den Kulturapfel sind weltweit bislang 51 mendelnde Merkmale kartiert und gelistet worden. In der Datenbank der Rosengewächse (https://www.rosaceae.org) sind insgesamt 121 genetische Karten mit Tausenden von molekularen Markern für den Kulturapfel und nahverwandte Wildarten der Gattung Malus gelistet. Darüber hinaus stehen den Züchtern und Züchtungsforschern verschiedene SNP-Marker-Arrays zur Verfügung, die in der Anzahl ihrer Marker von rund 8.000 (8k) bis 480.000 (480k) variieren. Solche Tools bieten enorme Möglichkeiten in der Genotypisierung, die von der Validierung einer Sortenechtheit, über die Erstellung bislang unbekannter Pedigrees, bis hin zu Genomweiten Assoziationsstudien und einer Genomweiten Selektion reichen. Erste Arbeiten dazu wurden bereits durchgeführt und die erzielten Ergebnisse sind vielversprechend (Jung et al., 2022).
Mehr als 200 genetische Karten existieren inzwischen auch für die unterschiedlichsten Prunus-Arten. Allein 51 davon sind für die Süßkirsche gelistet. Hier wurden bislang nur wenige Merkmale identifiziert, die den Vererbungsregeln Gregor Mendels folgen. Unter diesen ist vor allem die Fruchtgröße zu nennen, die für die Süßkirsche eines der wirtschaftlich bedeutendsten Merkmale darstellt. Für dieses Merkmal gibt es erste molekulare Marker, mit denen eine gezielte Auswahl von Kreuzungseltern möglich ist. Ein anderes bedeutendes mendelndes Merkmal der Süßkirsche ist die Selbstinkompatibilität. Die Erforschung der genetischen Ursachen für dieses Merkmal hat zur Entwicklung molekularer Marker und zur Identifizierung von mutierten (nicht funktionsfähigen) S-Allelen geführt, mit deren Hilfe es in den letzten Jahren gelungen ist, qualitativ hochwertige und selbstfertile Süßkirschensorten zu züchten.
Auch für Kirschen gibt es einen ersten SNP-Marker-Chip mit rund 15.000 (15k) SNP-Markern. Dieser ist für die Genotypisierung von Süßkirschen gut geeignet. Für andere Prunus-Arten, wie die Sauerkirsche ist dieser Array nicht optimal. Das liegt vor allem an der Komplexität des auto-allotetraploiden Genoms der Sauerkirsche und dem Fehlen hochwertiger Genomsequenzinformationen.
Solche hochwertigen Genomsequenzen, welche künftig die Identifikation von Kandidatengenen erleichtern werden, gibt es inzwischen für ausgewählte und ökonomisch bedeutende Apfelsorten, wie 'Golden Delicious' und 'Gala', aber auch für Wildarten wie M. baccata, M. prunifolia, M. sieversii und M. sylvestris, die eng mit der Evolution des Kulturapfels verbunden sind. Bei der Süßkirsche gibt es inzwischen auch eine erste Genomsequenz der Sorte 'Tieton'.
Rückblickend lässt sich feststellen, dass die meisten dieser Entwicklungen nicht, oder nicht so einfach, ohne die grundlegenden Erkenntnisse von Gregor Mendel möglich gewesen wären. Diese Erkenntnisse haben die Genetik der Kulturpflanzen auf eine höhere Stufe gestellt und ihr zahlreiche und bahnbrechende Möglichkeiten eröffnet.
Die Autorinnen und Autoren erklären, dass keine Interessenskonflikte vorliegen.
Alston, F.H., K.L. Phillips, K.M. Evans, 2000: A Malus gene list. Acta Horticulturae 538, 561-570, DOI: 10.17660/ActaHortic.2000.538.98.
Alston, F.H., J.B. Briggs, 1970: Inheritance of hypersensitivity to rosy apple aphid Dysaphis plantaginea in apple. Canadian Journal of Genetics and Cytology 12 (2), 257-258, DOI: 10.1139/g70-037.
Arber, W., S. Linn, 1969: DNA modification and restriction. Annual Review of Biochemistry 38, 467-500, DOI: 10.1146/annurev.bi.38.070169.002343.
Börner, C., F. Gollmick, 1943: Blutlausimmune Naumburger Edelapfelzüchtungen. Angewandte Botanik 25 (10), 144.
Bošković, R., K. Russell, K.R. Tobutt, 1997: Inheritance of stylar ribonucleases in cherry progenies, and reassignment of incompatibility alleles to two incompatibility groups. Euphytica 95, 221-228, DOI: 10.1023/A:1002945529157.
Bošković, R., K.R. Tobutt, 1996: Correlation of stylar ribonuclease zymograms with incompatibility alleles in sweet cherry. Euphytica 90, 245-250, DOI: 10.1007/BF00023865.
Bošković, R., K.R. Tobutt, 2001: Genotyping cherry cultivars assigned to incompatibility groups, by analysing stylar ribonucleases. Theoretical and Applied Genetics 103, 475-485, DOI: 10.1007/PL00002906.
Brady, N.C., 1991: Advances in agronomy. Academic Press.
Bramstedt, F., 1938: Der Nachweis der Blutlausunanfälligkeit der Apfelsorten auf histologischer Grundlage. (Aus der Biologischen Reichsanstalt für Land- und Forstwirtschaft, Zweigstelle Naumburg/Saale). Zeitschrift für Pflanzenkrankheiten (Pflanzenpathologie) und Pflanzenschutz 48 (9/11), 480-488.
Branscheidt, P., 1931: Weitere Mitteilungen über die Befruchtungsverhältnisse bei Obst, insbesondere bei Kirschen. Gartenbauwissenschaft 4, 387-427.
Brown, A.G., 1955: Incompatibility. John Innes Inst. Annu. Rpt. for 1954, 7-8.
Bus, V., A. White, S. Gardiner, R. Weskett, C. Ranatunga, A. Samy, M. Cook, E. Rikkerink, 2002: An update on apple scab resistance breeding in New Zealand. Acta Horticulturae 595, 43-47, DOI: 10.17660/ActaHortic.2002.595.5.
Bus, V.G. M.D. Chagné, H.C.M. Bassett, D. Bowatte, F. Calenge, J.M. Celton, C.E. Durel, M.T. Malone, A. Patocchi, A.C. Ranatunga, E.H.A. Rikkerink, D.S. Tustin, J. Zhou, S.E. Gardiner, 2008: Genome mapping of three major resistance genes to woolly apple aphid (Eriosoma lanigerum Hausm.). Tree Genetics and Genomes 4 (2), 223-236, DOI: 10.1007/s11295-007-0103-3.
Bus, V., E.H.A. Rikkerink, V. Caffier, C E. Durel, M. Plummers, 2011: Revision of nomenclature of the differential host-pathogen interactions of Venturia inaequalis and Malus. Annual Review of Phytopathology 49, 391-413, DOI: 10.1146/annurev-phyto-072910-095339.
Cai, Y., L. Wang, C.O. Ogutu, Q. Yang, B. Luo, L. Liao, B. Zheng, R. Zhang, Y. Han, 2021: The MADS-box gene PpPI is a key regulator of the double-flower trait in peach. Physiologia Plantarum 173 (4), 2119-2129, DOI: 10.1111/ppl.13561.
Cevik, V., G.J. King, 2002: High-resolution genetic analysis of the Sd-1 aphid resistance locus in Malus spp. Theoretical and Applied Genetics 105, 346-354, DOI: 10.1007/s00122-002-0904-6.
Chagne, D., C.M. Carlisle, C. Blond, R.K. Volz, C.J. Whitworth, N.C. Oraguzie, R.N. Crowhurst, A.C. Allan, R.V. Espley, R.P. Hellens, S.E. Gardiner, 2007: Mapping a candidate gene (MdMYB10) for red flesh and foliage colour in apple. BMC Genomics 8, 212, DOI: 10.1186/1471-2164-8-212.
Coen, E., E. Meyerowitz, 1991: The war of the whorls: genetic interactions controlling flower development. Nature 353, 31-37, DOI: 10.1038/353031a0.
Crandall, C.S., 1926: Apple breeding at the University of Illinois. Illinois Agricultural Experiment Station Bulletin 275, 341-600.
Crane, M.B., A.G. Brown, 1937: Incompatibility and sterility in the sweet cherry, Prunus avium L. Journal of Pomology and Horticultural Science 15, 86-116, DOI: 10.1080/03683621.1938.11513493.
Crane, M.B., J.C. Lawrence, 1929: Genetical and cytological aspects of incompatibility and sterility in cultivated fruits. Journal of Pomology and Horticultural Science 7, 276-301, DOI: 10.1080/03683621.1928.11513345.
Crane, M.B., R.M. Greenslade, A.M. Massee, H.M. Tybeman, 1936: Studies on the resistance and immunity of apples to the woolly aphis, Eriosoma lanigerum (Hausm.). Journal of Pomology and Horticultural Science 14 (2), 137-163, DOI: 10.1080/03683621.1937.11513467.
Cummins, J.N., H.S. Aldwinckle, 1983: Breeding apple rootstocks. Plant Breeding Reviews 1, 294-394, DOI: 10.1002/9781118060988.ch10.
de Nettancourt, D., 1977: Incompatibility in Angiosperms. Springer, Berlin.
de Vries, D.P., 1968: Compatibility of cherries in the Netherlands. Euphytica 17, 207-215, DOI: 10.1007/BF00021212.
Dirlewanger, E., E. Graziano, T. Joobeur, F. Garriga-Calderé, P. Cosson, W. Howad, P. Arús, 2004: Comparative mapping and marker-assisted selection in Rosaceae fruit crops. Proceedings of the National Academy of Science USA 101, 9891-9896, DOI: 10.1073/pnas.0307937101.
Dirlewanger, E., J. Claverie, A.F. Iezzoni, A. Wünsch, 2009: Sweet and sour cherries: linkage maps, QTL detection and marker assisted selection. In Genetics and genomics of Rosaceae, Springer, New York, NY, pp. 291-313.
Dochnahl, F.J., 1855: Der sichere Führer in der Obstkunde. Band 1: Aepfel. Wilhelm Schmid’s Buch- und Kunsthandlung, Nürnberg.
Emeriewen, O.F., K. Richter, S. Piazza, D. Micheletti, G.A.L. Broggini, T. Berner, J. Keilwagen, M.-V. Hanke, M. Malnoy, A. Peil, 2018. Towards map-based cloning of FB_Mfu10: identification of a receptor-like kinase candidate gene underlying the Malus fusca fire blight resistance locus on linkage group 10. Molecular Breeding 38, 106, DOI: 10.1007/s11032-018-0863-5.
Emeriewen, O.F., A. Peil, K. Richter, E. Zini, M.-V. Hanke, M. Malnoy, 2017: Fire blight resistance of Malus ×arnoldiana is controlled by a quantitative trait locus located at the distal end of linkage group 12. European Journal of Plant Pathology 148 (4), 1011-1018, DOI: 10.1007/s10658-017-1152-6.
Emeriewen, O.F., K. Richter, H. Flachowsky, M. Malnoy, A. Peil, 2021: Genetic analysis and fine mapping of the fire blight resistance locus of Malus xarnoldiana on linkage group 12 reveal first candidate genes. Frontiers in Plant Science 12, 667133, DOI: 10.3389/fpls.2021.667133.
Espley, R.V., C. Brendolise, D. Chagne, S. Kutty-Amma, S. Green, R. Volz, J. Putterill, H.J. Schouten, S.E. Gardiner, R.P. Hellens, 2009: Multiple repeats of a promoter segment causes transcription factor autoregulation in red apples. The Plant Cell 21 (1), 168-183, DOI: 10.1105/tpc.108.059329.
Fahrentrapp, J., G.A.L. Broggini, M. Kellerhals, A. Peil, K. Richter, E. Zini, C. Gessler, 2013: A candidate gene for fire blight resistance in Malus × robusta 5 is coding for a CC-NBS-LRR. Tree Genetics and Genomes 9 (1), 237-251, DOI: 10.1007/s11295-012-0550-3.
Flachowsky, H., R. Töpfer, 2021: Obst- und Rebenzüchtung im Zeitraffer. Journal für Kulturpflanzen. 73 (7-8), 197-203, DOI: 10.5073/JfK.2021.07-08.03.
Friedli, M., A. Häseli, P. Stefani, 2021: Biokernobst, Sortenliste 2021, Nr. 1451, URL: https://www.fibl.org/fileadmin/documents/shop/1451-biokernobst.pdf.
Gao, Z.S., W.E. van de Weg, 2006: The V-f gene for scab resistance in apple is linked to sub-lethal genes. Euphytica 151 (1), 123-133, DOI: 10.1007/s10681-005-9082-3.
Gardiner, S., H. Bassett, J. Murdoch, S. Meech, M. Cook, V. Bus, C. Ranatunga, E. Rikkerink, 2001: Major pest and disease resistance loci in apple available to breeders. In: Plant and Animal Genome, IX Conf., San Diego, CA, USA. URL: http://www.intl- pag.org/pag/9/abstracts/P5h_13.html.
Gardiner, S.E., V.G.M. Bus, R.L. Rusholme, D. Chagné, E H.A. Rikkerink, 2007: Apple. In: Fruits and Nuts (Volume 4), Genome Mapping and Molecular Breeding, 1-62, Springer-Verlag, DOI: 10.1007/978-3-540-34533-6_1.
Giliomee, J.H., D.K. Strydom, H.J. Van Zyl, 1968: Northern Spy, Merton and Malling–Merton rootstocks susceptible to woolly aphid, Eriosoma lanigerum, in the Western Cape. South African Journal of Agricultural Science 11, 183-186.
Hanke, M.-V., H. Flachowsky, A. Peil, O.F. Emeriewen, 2020: Malus × domestica apple. In Biotechnology of fruit and nut crops. 2nd Edition. Litz, R., F. Pliego-Alfaro, J.I. Hormaza (Hrsg), Cambridge: CAB International), 440-473.
Hanke, M.-V., H. Flachowsky, A. Peil, C. Hättasch, 2007: No flower no fruit: genetic potentials to trigger flowering in fruit trees. Genes, Genomes and Genomics 1 (1), 1-20.
Hauagge, R., J.N. Cummins, 1991: Genetics of length of dormancy period in Malus vegetative buds. Journal of the American Society for Horticultural Science 116 (1), 121-126, DOI: 10.21273/JASHS.116.1.121.
Hossfeld, U., M.V. Simunek, M. Mielewczik, 2017: Die „Wiederentdeckung“ der Mendelschen Gesetze im Kontext neuer Forschungen. Nova Acta Leopoldina NF 413, 135-153.
Hough, L.F., J.R. Shay, D.F. Dayton, 1953: Apple scab resistance from Malus floribunda Sieb. Proceedings of the American Society for Horticultural Science 62, 341-347.
Irish, V., 2017: The ABC model of floral development. Current Biology 27 (17), R887-R890, DOI: 10.1016/j.cub.2017.03.045.
Jancke, O., 1937: Über die Blutlausanfälligkeit von Apfelsorten, wilden Malusarten und -bastarden sowie die Züchtung blutlausfester Edeläpfel und Unterlagen. Phytopathologische Zeitschrift 10, 185.
Jung, M., M. Roth, M. José Aranzana, A. Auwerkerken, M. Bink, C. Denancé, C. Dujak, C.-E. Durel, C. Font i Forcada, C. Cantin, W. Guerra, N.P. Howard, B. Keller, M. Lewandowski, M. Ordidge, M. Rymenants, N. Sanin, B. Studer, E. Zurawicz, F. Laurens, A. Patocchi, H. Muranty, 2022: The apple REFPOP – a reference population for genomics-assisted breeding in apple. Horticulture Research 7, 189, DOI: 10.1038/s41438-020-00408-8.
King, G.J., F.H. Alston, I. Battle, E. Chevreau, C. Gessler, J. Janse, P. Lindhout, A.G. Manganaris, S. Sansavini, H. Schmidt, K.R. Tobutt, 1991: The ‘European apple genome mapping project’— developing a strategy for mapping genes coding for agronomic characters in tree species. Euphytica 56, 89-94, DOI: 10.1007/BF00041748.
Klocko, A.L., E. Borejsza-Wysocka, A.M. Brunner, O. Shevchenko, H. Aldwinckle, S.H. Strauss, 2016: Transgenic Suppression of AGAMOUS genes in apple reduces fertility and increases floral attractiveness. PLoSOne 11 (8), e0159421, DOI: 1371/journal.pone.0159421.
Knight, R.L., 1969: Abstract bibliography of plant breeding and genetics to 1965, Prunus. Eastern Press Ltd., London, 540-544.
Knight, R.L., J.B. Briggs, A.M. Massee, H.N. Tydeman, 1962: The inheritance of resistance to woolly aphid, Eriosoma lanigerum (Hausm.), in the apple. Journal of Horticultural Science 37, 207-218.
Kobel, F., 1927: Untersuchungen über die Blüh- und Fruchtbarkeitsverhältnisse bei Obstbäumen. Die Gartenwelt 31, 649-651.
Kobel, F., P. Steinegger, J. Anliker, 1938: Weitere Untersuchungen über die Befruchtungsverhältnisse der Kirschsorten. Landwirtschaftliches Jahrbuch der Schweiz 52, 564-895.
Lane, D., H. Schmid, 1984: Lapins and Sunburst sweet cherry. Canadian Journal of Plant Science 64, 211-214, DOI: 10.4141/cjps84-029.
Lapins, K.O., 1971: Stella, a self-fruitful sweet cherry. Canadian Journal of Plant Science 51, 252-253, DOI: 10.4141/cjps71-051.
Lempe, J., A. Peil, H. Flachowsky, 2022: Time-resolved analysis of candidate gene expression and ambient temperature during bud dormancy in apple. Frontiers in Plant Science 12, 803341, DOIi: 10.3389/fpls.2021.803341.
Lewis, D., L.K. Crowe, 1954: Structure of the Incompatibility Gene. IV. Types of Mutation in Prunus avium L. Heredity 8, 357-363.
Maxam, A.M., W. Gilbert, 1977: A new method for sequencing DNA. Proceedings of the National Academy of Science USA 74 (2), 560-564, DOI: 10.1073/pnas.74.2.560.
Mendel, G., 1866: Versuche über Pflanzenhybriden. Verhandlungen des Naturforschenden Vereines in Brünn, Bd. IV, 3-47.
Mihatsch, H., G. Schumann, 1971: Untersuchungen über Befruchtungsverhältnisse bei Süßkirsche (Prunus avium L.). Archiv für Gartenbau 19, 373-378.
Morgan, T.H., 1910: Sex limited inheritance in Drosophila. Science, 120-122, DOI: 10.1126/science.32.812.120.
Moser, M., E. Asquini, G.V. Miolli, K. Weigl, M.-V. Hanke, H. Flachowsky, A. Si-Ammour, 2020: The MADS-box gene MdDAM1 controls growth cessation and bud dormancy in apple. Frontiers in Plant Science 11, 1003, DOI: 10.3389/fpls.2020.01003.
Mullis, K.B., F. Faloona, S. Scharf, R. Saiki, G. Horn, H. Erlich, 1986: Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harbor Symposium on Quantitative Biology 51 (1), 263-273.
Nirenberg, M.W., H.J. Matthaei, 1961: The Dependence of cell- free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proceedings of the National Academy of Sciences of the United States of America 47(10), 1588-1602, DOI: 10.1073/pnas.47.10.1588.
Olmstead, J.W., A.M. Sebolt, A. Cabrera, S.S. Sooriyapathirana, S. Hammar, G. Iriarte, D. Wang, C.Y. Chen, E. van der Knaap, A.F. Iezzoni, 2008: Construction of an intra-specific sweet cherry (Prunus avium L.) genetic linkage map and synteny analysis with the Prunus reference map. Trees Genetics and Genomes 4, 897-910, DOI: 10.1007/s11295-008-0161-1.
Orel, V., M. Vavra, 1968: Mendel's program for the hybridization of apple trees. Journal of the History of Biology 1 (2), 219-224, DOI: 10.1007/BF00351921.
Otto, D., R. Petersen, B. Brauksiepe, P. Braun, E.R. Schmidt, 2014: The columnar mutation (“Co gene”) of apple (Malus × domestica) is associated with an integration of a Gypsy-like retrotransposon. Molecular Breeding 33, 863-880, DOI: 10.1007/s11032-013-0001-3.
Parravicini, G., C. Gessler, C. Denancé, P. Lasserre-Zuber, E. Vergne, M.-N. Brisset, A. Patocchi, C.-E. Durel, G.A.L. Broggini, 2011: Identification of serine/threonine kinase and nucleotide-binding site–leucine-rich repeat (NBS-LRR) genes in the fire blight resistance quantitative trait locus of apple cultivar ‘Evereste’. Molecular Plant Pathology 12, 493-505, DOI: 10.1111/j.1364-3703.2010.00690.x.
Pascal, T., R. Aberlenc, C. Confolent, M. Hoerter, E. Lecerf, C. Tuéro, P. Lambert, 2017: Mapping of new resistance (Vr2, Rm1) and ornamental (Di2, pl) Mendelian trait loci in peach. Euphytica 213, 1-12, DOI: 10.1007/s10681-017-1921-5.
Peil, A., O.F. Emeriewen, A. Khan, S. Kostick, M. Malnoy, 2020: Status of fire blight resistance breeding in Malus. Journal of Plant Pathology 103, 3-12, DOI: 10.1007/s42161-020-00581-8.
Peil, A., O.F. Emeriewen, K. Richter, T. Wöhner, M.,Malnoy, M.-V. Hanke, H. Flachowsky, 2014: Vergleichende genetische Kartierung der Feuerbrandresistenz bei Malus sp.. Journal für Kulturpflanzen 66 (12), 409-416, DOI: 10.5073/JfK.2014.12.02.
Peil, A., T. Garcia-Libreros, K. Richter, F.C. Trognitz, B. Trognitz, M.-V. Hanke, H. Flachowsky, 2007: Strong evidence for a fire blight resistance gene of Malus robusta located on linkage group 3. Plant Breeding 126, 270-475, DOI: 10.1111/j. 1439-0523.2007.01408.x.
Picard, C., T. Afonso, A. Benko‐Beloglavec, O. Karadjova, S. Matthews‐Berry, S.A. Paunovic, M. Pietsch, P. Reed, D.J. Van Der Gaag, M. Ward, 2018: Recommended regulated non‐quarantine pests (RNQPs), associated thresholds and risk management measures in the European and Mediterranean region. EPPO Bulletin 48 (3), 552-568, DOI: 10.1111/epp.12500.
Pijut, P.M., 2020: Prunus Spp. Cherry. In: Biotechnology of fruit and nut crops. Litz, R., F. Pliego-Alfaro, J.I. Hormaza (Hrsg) 2nd Edition. Cambridge: CAB International, 440-473.
Ritter Beck von Mannagetta, G., F. Abel, 1894: Sprechabende über das Gesammtgebiet der Horticultur in Wien. Wiener illustrirte Garten-Zeitung, Jahrgang 19, 227-228.
Rock, G., D. Zeiger, 1974: Woolly apple aphid infests Malling and Malling–Merton rootstocks in propagation beds in North Carolina. Journal of Economic Entomology 67, 137-138, DOI: 10.1093/jee/67.1.137a.
Rosyara, U.R., M.C. Bink, E. van de Weg, G. Zhang, D. Wang, A. Sebolt, E. Dirlewanger, J. Quero-Garcia, M. Schuster, A.F. Iezzoni, 2013: Fruit size QTL identification and the prediction of parental QTL genotypes and breeding values in multiple pedigreed populations of sweet cherry. Molecular Breeding, 32 (4), 875-887, DOI: 10.1007/s11032-013-9916-y.
Sanger, F., S. Nicklen, A.R. Coulson, 1977: DNA sequencing with chain-terminating inhibitors. Proceedings of the National Academy of Sciences 74, 5463-5467.
Schmidt, H., B. Wolfram, R. Boškovć, 1999: Befruchtungsverhältnisse bei Süßkirschen. Erwerbsobstbau 41, 42-45.
Schröpfer, S., J. Lempe, O.F. Emeriewen, H. Flachowsky, 2022: Recent Developments and Strategies for the Application of Agrobacterium-mediated transformation of apple Malus × domestica Borkh. Frontiers in Plant Science 13, 928292, DOI: 10.3389/fpls.2022.928292.
Schuster, M., 2020: Self-incompatibility (S) genotypes of cultivated sweet cherries – An overview update 2020. Data from OpenAgrar Repository, DOI: 10.5073/20201016-141600.
Sen Gupta, G.C., P.W. Miles, 1975: Studies on the susceptibility of varieties of apple to the feeding of two strains of woolly aphis (Homoptera) in relation to the chemical content of the tissues of the host. Austin Journal of Agricultural Research 26, 157-168.
Shay, R., 1962: Apple. Resistance to diseases and insects. Handbuch der Pflanzenzüchtung 6, 668-671.
Sonneveld, T., T.P. Robbins, R. Bošković, K.R. Tobutt, 2001: Cloning of six cherry self-incompatibility alleles and development of allele-specific PCR detection. Theoretical and Applied Genetics 102, 1046-1055, DOI: 10.1007/s001220000525.
Sonneveld, T., K.R. Tobutt, T.P. Robbins, 2003: Allele-specific PCR detection of sweet cherry self-incompatibility (S) alleles S1 to S16 using consensus and allele-specific primers. Theoretical and Applied Genetics 107, 1059-1070, DOI: 10.1007/s00122-003-1274-4.
Sonneveld, T., T.P. Robbins, K.R. Tobutt, 2006: Improved discrimination of self-incompatibility S-RNase alleles in cherry and high throughput genotyping by automated sizing of first intron polymerase chain reaction products. Plant Breeding 125, 305-307, DOI: 10.1111/j.1439-0523.2006.01205.x.
Speyer, W., 1924: Über die Blutlausanfälligkeit von Apfelsorten. Angewandte Botanik 6,168-171.
Tanaka, N., M. Wada, S. Komori, H. Bessho, A. Suzuki, 2007: Functional analysis of MdPI, the PISTILLATA gene homologue of apple, in Arabidopsis. Journal of the Japanese Society for Horticultural Science 76, 125-132, DOI: 10.2503/jjshs.76.125.
Tao, R., H. Yamane, A. Sugiura, H. Murayama, H. Sassa, H. Mori, 1999: Molecular typing of S-alleles through identification, characterization and cDNA cloning for S-RNases in sweet cherry. Journal of the American Society of Horticultural Science 124, 224-233, DOI: 10.21273/JASHS.124.3.224.
Tehrani, G., S.K. Brown, 1992: Pollen-incompatibility and self-fertility in sweet cherry. In: Plant Breeding Reviews. Janick, J. (Hrsg), 9, 367-388, DOI: 10.1002/9780470650363.ch10.
Tobutt, K.R., 1994: Combining apetalous parthenocarpy with columnar growth habit in apple. Euphytica 77, 51-54, DOI: 10.1007/BF02551460.
Ushijima, K., H. Sassa, R. Tao, H. Yamane, A.M. Dandekar, T.M. Gradziel, H. Hirano, 1998: Cloning and characterization of cDNAs encoding S-RNases from almond (Prunus dulcis): primary structural features and sequence diversity of the S-RNases in Rosaceae. Molecular and General Genetics 260, 261-268, DOI: 10.1007/s004380050894.
Vahl v., E., 1965: Prüfung der Befruchtungsverhältnisse bei Süßkirschkreuzungen. Erwerbsobstbau 7, 126-129.
Vavra, M., V. Orel, 1971: Hybridization of pear varieties by Gregor Mendel. Euphytica 20, 60-67, DOI: 10.1007/BF00146775.
Volz, R.K., T.K. McGhie, S. Kumar, 2014: Variation and genetic parameters of fruit colour and polyphenol composition in an apple seedling population segregating for red leaf. Tree Genetics and Genomes 10 (4), 953-964, DOI: 10.1007/s11295-014-0734-0.
Wang, L., B. Yu, Y. Zhao, Y. Li, J. Guo, Y. Zhu, 2021: A putative 2OG-Fe (II) oxygenase’s response to gibberellin deficiency is related to the internodal growth of columnar apples. Acta Physiologiae Plantarum 43 (5), 1-11, DOI: 10.1007/s11738-021-03243-z.
Watson, J., F. Crick, 1953: Molecular structure of nucleic acids: a structure for deoxyribose nucleic acid. Nature 171, 737-738, DOI: 10.1038/171737a0.
Weigel, D., E.M. Meyerowitz, 1994: The ABCs of floral homeotic genes. Cell 78 (2), 203-209, DOI: 10.1016/0092-8674(94)90291-7.
Wiersma, P.A., Z. Wu, L. Zhou, C. Hampson, F. Kappel, 2001: Identification of new self-incompatibility alleles in sweet cherry (Prunus avium L.) and clarification of incompatibility groups by PCR and sequencing analysis. Theoretical and Applied Genetics 102, 700-708, DOI: 10.1007/s001220051700.
Wolters, P.J., H.J. Schouten, R. Velasco, A. Si-Ammour, P. Baldi, 2013: Evidence for regulation of columnar habit in apple by a putative 2OG-Fe(II) oxygenase. New Phytologist 200, 993-999, DOI: 10.1111/nph.12580.
Yamane, H.R., H.M. Tao, A. Sugiura, 2003: Identification of a non-S RNase, a possible ancestral form of S-RNases, in Prunus. Molecular Genetics and Genomics 269, 90-100, DOI: 10.1007/s00438-003-0815-5.
Yao, J.-L., Y.-H. Dong, B.A. Morris, 2001: Parthenocarpic apple fruit production conferred by transposon insertion mutations in a MADS-box transcription factor. Proceedings of the National Academy of Sciences 98, 1306-1311, DOI: 10.1073/pnas.98.3.1306.
Zhang, G., A.M. Sebolt, S.S. Sooriyapathirana, D. Wang, M.C.A.M. Bink, J.W. Olmstead, A. Iezzoni, 2010: Fruit size QTL analysis of an F1 population derived from a cross between a domesticated sweet cherry cultivar and a wild forest sweet cherry. Tree Genetics and Genomes 6, 25-36, DOI: 10.1007/s11295-009-0225-x.

 Suchen
Suchen