
Journal für Kulturpflanzen, 74 (11-12). S. 233–241, 2022 | DOI: 10.5073/JfK.2022.11-12.03 | Uptmoor und Kirchgesser
Mendel und der Beginn der modernen Genetik: zu gut um wahr zu sein?
Mendel and the onset of modern genetics: too good to be true?
 | Der Autor/Die Autorin 2022 Dies ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Namensnennung 4.0 International Lizenz (CC BY 4.0) zur Verfügung gestellt wird (https://creativecommons.org/licenses/by/4.0/deed.de). |
Zur Veröffentlichung eingereicht/angenommen: 29. Juli 2022/4. November 2022 |
Im 18. und 19. Jahrhundert wurden sowohl in Europa als auch in den USA Kreuzungsexperimente an Mais und Zierpflanzen, aber auch an der Erbse durchgeführt. Mendel kannte zumindest einen Teil der Literatur, als er seine „Versuche über Pflanzenhybriden“ 1866 veröffentlichte. Im Gegensatz zu seinen Vorgängern fielen Mendel die konstanten Zahlenverhältnisse auf, die in den Nachkommen seiner Kreuzungsexperimente zu beobachten waren. Nachdem die Arbeiten Mendels über mehrere Jahrzehnte weitgehend unbeachtet blieben, wurden seine Theorien zur Vererbungslehre 1900 von de Vries, Correns und Tschermak wiederentdeckt und experimentell bestätigt. Kurz darauf kamen erste Zweifel auf, da die Ergebnisse Mendels fast durchweg sehr nahe an seinen Erwartungswerten lagen. Ronald A. Fisher publizierte 1936 einen Artikel, der schlussfolgerte, dass ein Großteil von Mendels Ergebnissen zugunsten der theoretischen Spaltungsverhältnisse gefälscht worden sein müsste. Damit war die Mendel-Fisher-Kontroverse geboren. Zum Jubiläum der Veröffentlichung der Grundlagen der Vererbungslehre von Gregor Mendel brachten Stern & Sherwood 1966 ein Buch heraus, welches sowohl Mendels Texte auch die der Wiederentdeckung seiner Lehren sowie den kritischen Text von Fisher enthielten. Damit nahm die Mendel-Fisher-Kontroverse an Fahrt auf und sorgte für zahlreiche weitere Publikationen. Ob Mendel einfach Glück hatte, ob er wissentlich Pflanzen aussortierte, die nicht seinen Erwartungen entsprachen oder ob er einen Assistenten hatte, der zu gut wusste, was Mendel erwartete – wie Fisher vermutete – werden wir nicht mit letzter Sicherheit herausfinden können. Gesichert ist, dass die Mendelschen Vererbungsregeln bis heute ihre Gültigkeit haben und in Zukunft behalten werden.
Gregor Mendel, Ronald A. Fisher, Vererbungsregeln, Spaltungsverhältnisse, Erbse
In the 18th and 19th century crossing experiments using maize, ornamentals and peas were carried out in Europe and in the USA. Mendel knew at least a part of the literature when he published his famous paper "Experiments on Plant Hybridisation" with the original German title "Versuche über Pflanzenhybriden" in 1866. In contrast to his predecessors, Mendel recognised the constant segregation ratios appearing in the progenies of the crossings. After being unnoticed for decades, his fundamental theory of heredity was rediscovered and experimentally verified by de Vries, Correns and Tschermak in 1900. Shortly after the English translation of Mendel's work became available in 1901, Raphael Weldon, one of the founders of biometry, published his doubts on Mendel's data since all observed segregation ratios were extremely close to the expected ones. Ronald A. Fisher concluded in a paper from 1936 that a large part of Mendel's results must have been falsified to agree closely with the postulated segregation ratios. The Mendel-Fisher controversy was born. In 1966, 100 years after Mendel's original paper was released, Stern & Sherwood published a book comprising translations of Mendel's laws of inheritance, the rediscovery by de Vries and Correns and Fisher's paper. Subsequently, this led to the publication of numerous articles from different scientific disciplines on the Mendel-Fisher controversy. It may never be revealed, if it was simple luck, if Mendel sorted out plants, which did not represent the theory, or if he had an assistant "who knew too well what was expected", as Fisher speculated in 1936. Nonetheless, Mendel's work doubtlessly culminated in one of the most important breakthrough discoveries of the 19th century and is the starting point of modern genetics and plant breeding.
Gregor Mendel, Ronald A. Fisher, laws of inheritance, segregation ratios, pea
Mendel war nicht der erste, der mit Erbsen experimentierte und Kreuzungen durchführte. Thomas Andrew Knight bestäubte 1799 Erbsen mit weißen Samen mit dem Pollen von solchen mit grauen Samen und entdeckte, dass alle Nachkommen nur graue Samen hervorbrachten. Er machte Rückkreuzungen, bei denen er feststellte, dass wieder weißsamige Erbsen auftraten (Knight, 1799). Alexander Seton (1824, nach Zirkle, 1951) kreuzte grünsamigige mit weißsamigen Erbsen und stellte ebenfalls fest, dass alle Nachkommen eine identisch grüne Samenfarbe hatten. Nachdem er die grünen Samen wieder ausgesät hatte, konnte er Erbsensamen beider Farben ernten und bemerkte, dass keiner der Samen farblich intermediär war. Im selben Jahr fand John Goss (1824, nach Zirkle, 1951), dass in einer Kreuzung von Erbsenpflanzen mit blauen und weißen Erbsen in der ersten Generation nur weiße Samen auftraten, in der zweiten Generation fand er Nachkommen die nur weiße, nur blaue oder sowohl weiße als auch blaue Samen in denselben Hülsen hatten. In der dritten Generation produzierten die blausamigen Erbsen nur blausamige Nachkommen, während die weißsamigen Pflanzen entweder nur weiße, nur blaue oder wieder beide Samenfarben in ihren Hülsen trugen. Knight (1824) kommentierte die Arbeiten und machte die richtigen Aussagen über Dominanz und Rezessivität, aber er zählte die jeweiligen Anteile nicht und konnte deshalb keine eindeutigen Regeln ableiten (Zirkle, 1951).
Erbsen erfreuten sich in der jüngeren Neuzeit großer Beliebtheit als Gemüsekultur. So schrieb Marie de Rabutin-Chantal vom Hof Louis XIV, in dessen Küchengarten in Versailles Erbsen kultiviert wurden: „Das Erbsenkapitel dauert an: die Ungeduld, sie zu essen, das Vergnügen, sie gegessen zu haben, und die Freude, sie wieder zu essen – das sind die drei Themen, über die man seit vier Tagen redet.“ Der spätere US-Präsident Thomas Jefferson baute Erbsen in seinem Garten in Virginia an und notierte am 26. Februar 1778 in sein Gartenbuch „sowed a patch of Hotspur peas“. Hotspurs sind besonders schnellwüchsige Sorten (Woolridge, 1677). Thomas Andrew Knight erstellte im frühen 19. Jahrhundert die ersten gezielten Erbsenzüchtungen. Aus den Sorten ‘Knight’s Tall Green Marrow’ und ‘Knight’s Dwarf Green Marrow’ sind im Laufe des 19. Jahrhunderts viele weitere Sorten entstanden. Ziel der damaligen Züchter war es, die Erträge zu steigern sowie hochwachsende Sorten, die Rankhilfen benötigen und kleine, runde Samen hatten, durch solche mit niedrigem Wuchs und großen runzligen Samen zu ersetzen (Hedrick et al., 1928). Es gab zwar bereits vorher Landsorten mit niedrigem Wuchs sowie Marrowfats mit großen, runzligen Samen (Townsend, 1725), allerdings dürften die meisten Sorten hochwachsend und rundsamig gewesen sein. John Woolridge schrieb 1677, dass die Erbse schon lange in England angebaut werde, die süßen und delikaten Sorten aber erst spät eingeführt worden seien. Im Gegensatz zu den rundsamigen Palerbsen entsprechen Marrowfats den heutigen Markerbsen. Der Zuckergehalt von Markerbsen ist mit maximal 7 bis 9 % höher als der von Palerbsen (4 bis 5 %), woraus ein deutlich süßerer Geschmack resultiert. Der höhere Zuckergehalt der Markerbse geht zunächst mit einem höheren Wassergehalt einher. Das führt nach dem Wasserentzug zum typisch runzligen Erscheinungsbild reifer Samen (wrinkled seeded peas). Tracy listet bereits 1903 für das U.S. Department of Agriculture ca. 500 Sorten, darunter normalwüchsige, Halbzwerge und Zwerge mit sehr niedrigem Wuchs, Marrowfats mit runzligen Samen, Sorten mit runden Samen für die Konservenindustrie und Zuckererbsen auf.
Obwohl Mendel (1866) z. B. die unten genannte Publikation zur Hybridisierung bei Pflanzen von Gärtner (1849) zitierte, ist nicht überliefert, ob er mit den früheren Arbeiten speziell an Erbsen vertraut war. Mit der Erbse stand ihm allerdings eine Modellpflanze zur Verfügung, die eine große Variation in vielen einfach zu bonitierenden Eigenschaften wie Farbe der Samenschale und Kotyledonen, Samenform, Farbe und Form der Hülsen, Wuchshöhe und Blütenfarbe aufwies. Die Erbse ist ein Selbstbefruchter mit 2n=14 Chromosomen und einem ausgesprochen großen Genom von 4,45 Gb (Kreplak et al., 2019). Obwohl sowohl bei den Wildformen als auch bei der Kulturerbse eine Bestäubung über Insekten möglich ist, herrscht bei den im Anbau befindlichen Sorten Kleistogamie vor. Die Selbstbestäubung findet demnach in der geschlossenen Blüte statt. Mendel entschied sich auch aufgrund der Kleistogamie für die Erbse als Modellpflanze, da Selbstungen, also Inzucht durch Selbstbestäubungen, leicht und sicher vonstattengehen. Mendel schreibt 1866: „Versuche (…) führten zu dem Resultate, dass das Genus Pisum den gestellten Anforderungen hinreichend entspreche.“ Die künstliche Befruchtung, bei der er die Knospe zum Entfernen der noch unreifen Staubfäden mit der Pinzette öffnete, um die Narbe später mit fremdem Pollen zu bestäuben, beschreibt er zwar als umständlich aber zumeist gelingend (Mendel, 1866). Schon in seinen einleitenden Bemerkungen erwähnt Mendel (1866) die Regelmäßigkeit, die bei Hybriden zu beobachten ist, wenn Zierpflanzen zur Erzeugung neuer Farbvarianten künstlich befruchtet werden. Auch zitiert er die Arbeiten von Gärtner (1849) über die Erzeugung von Hybriden bei Pflanzen. Gärtner wiederum zitierte die Kreuzungsexperimente, die Knight (1799) an Erbsen durchgeführt hatte (Zirkle, 1951). Damit ist also sicher, dass Gregor Mendel Studien zur Hybridisierung kannte und nicht völlig unvoreingenommen seine Experimente begann.
Von den vielen Merkmalen, die Mendel untersuchte, sind Form und Farbe der Samen bzw. Kotelydonen die wohl bekanntesten (Mendel, 1866; Miko, 2008). Zunächst arbeitete er mit reinerbigen Pflanzen, die er über mehrere Generationen geselbstet hatte. Danach kreuzte er Elternlinien mit verschiedenen Eigenschaften untereinander. Obwohl bereits vorher gegenteiliges beobachtet wurde (Knight, 1799; 1824; Gärtner, 1849), ging man weitestgehend davon aus, dass Nachkommen immer einer Mischung aus ihren Eltern entsprechen, was einem intermediären Erbgang gleichkommt. Die Nachkommen von Elternlinien mit runden und eckigen Samen konnten diese Annahmen aber nicht stützen, denn alle waren rund und folgten einem dominanten Erbgang. Da alle Nachkommen in der ersten Filialgeneration (F1) gleich aussahen, formulierte Mendel daraus die Uniformitätsregel, das 1. Mendelsche Gesetz.
Als Mendel in einem weiteren Schritt die F1-Pflanzen, die genetisch alle identisch waren, mit sich selbst kreuzte, traten sowohl runde als auch eckige Samen im Verhältnis 3:1 auf. Mendel stellte fest, dass Eigenschaften, die in der F1 nicht mehr auftauchten, weiterhin rezessiv vorhanden waren und an spätere Generationen vererbt werden konnten. Er nutzte dabei die Denomination AA für den Elter mit runden Samen und aa für den Elter mit eckigen Samen. In der F1 kamen nur Aa Genotypen mit der dominanten Eigenschaft der runden Samenform vor. In der F2 glich ein Viertel der Genotypen wieder dem dominanten Elter AA, ein weiteres Viertel dem rezessiven Elter aa, die Hälfte blieb genotypisch identisch mit der F1-Generation (Aa). Da letztere phänotypisch nicht vom reinerbigen Genotyp AA unterschieden werden konnten, kam das Spaltungsverhältnis 3:1 zustande. Heute wissen wir, dass A und a den zwei Allelen eines Gens entsprechen. Mendel (1866) formulierte aus seinen Ergebnissen die Spaltungsregel. Er sah in seinen Ergebnissen, Gärtner (1849) bestätigt, demzufolge „Hybriden die Neigung besitzen, zu den Stammarten zurückzukehren“ (Mendel 1866), also bei Selbstung oder Kreuzung untereinander aufspalten.
Um herauszufinden, was passiert, wenn man Eltern miteinander kreuzt, die sich in zwei Eigenschaften voneinander unterscheiden, kreuzte Mendel Pflanzen mit rund-gelben Samen (AABB) mit solchen mit kantig-grünen Samen (aabb). Der Begriff „kantig“ ist hierbei mit dem oben genannten Begriff „runzelig“ für Markerbsen gleichzusetzen. Mendel (1866) benutzte beide Begriffe. In der F1-Generation waren alle Nachkommen rund und gelb (AaBb), da das die dominanten Eigenschaften waren. In der F2-Generation traten alle möglichen Kombinationen der Merkmale auf: rund-gelbe (AABB, AABb, AaBB, AaBb), kantig-gelbe (aaBB, aaBb), rund-grüne (AAbb, Aabb) und kantig-grüne (aabb) Erbsensamen. Für sich genommen traten die Eigenschaften wieder im Verhältnis 3:1 auf, in oben genannter Kombination allerdings im Verhältnis 9:3:3:1. Mendel (1866) erwähnte dieses Spaltungsverhältnis nicht. Er baute die Samen der F2 Generation wieder an und stellte fest, dass die Pflanzen aus kantig-grünen Samen in der Folgegeneration konstant blieben. Die Pflanzen, die aus rund-gelben Samen gezogen wurden, erzeugten im Verhältnis 1:2:2:4 nur rund-gelbe Samen, rund-gelbe und -grüne Samen, rund- und kantig-grüne Samen sowie alle vier Varianten der F2. Es gab also auch hier wieder die rund-gelbe homozygote Ausgangsform, während ein Teil homozygot für das jeweils dominante Merkmal geworden war und ein weiterer Teil sich äquivalent zur F1-Hybride verhielt. Mendel (1866) bewies mit diesen Versuchen, dass die Merkmale unabhängig voneinander vererbt wurden, woraus sich die Unabhängigkeitsregel ableitet.
Mendel hat bei seinen Arbeiten nicht nur die richtigen Eigenschaften gewählt, nämlich solche, die über einzelne Gene vererbt werden, er hatte auch Glück, denn die Gene für die beiden oben genannten Eigenschaften liegen tatsächlich auf unterschiedlichen Chromosomen. Dazu kommt, dass die bekanntesten Merkmale die er untersuchte – Samenform und -farbe – schon auf der Mutterpflanze beobachtet werden konnten. Sein größter Verdienst war es – im Gegensatz zu seinen Vorgängern – exakt zu zählen und mit großen Zahlen zu arbeiten, klare Hypothesen zu formulieren und diese anhand seiner Erwartungen zu überprüfen. So untersuchte er in seinem Experiment zur Ableitung der Spaltungsregel mehr als 8.023 Samen von 258 Pflanzen. Davon waren 6022 gelb und 2001 grün. Gelb trat also zu grün im Verhältnis 3,01:1 auf, was dem theoretischen Spaltungsverhältnis von 3:1 schon sehr nahekam. Mit ähnlich hohen Zahlen arbeitete er, als er die Samenform untersuchte (Abb.). Das Spaltungsverhältnis 9:3:3:1 zur Unabhängigkeitsregel basiert auf 556 Samen von 15 Pflanzen, die er im Folgejahr alle zur Aussaat brachte und von denen er abzüglich der Verluste 529 wieder untersuchte (Mendel, 1866).
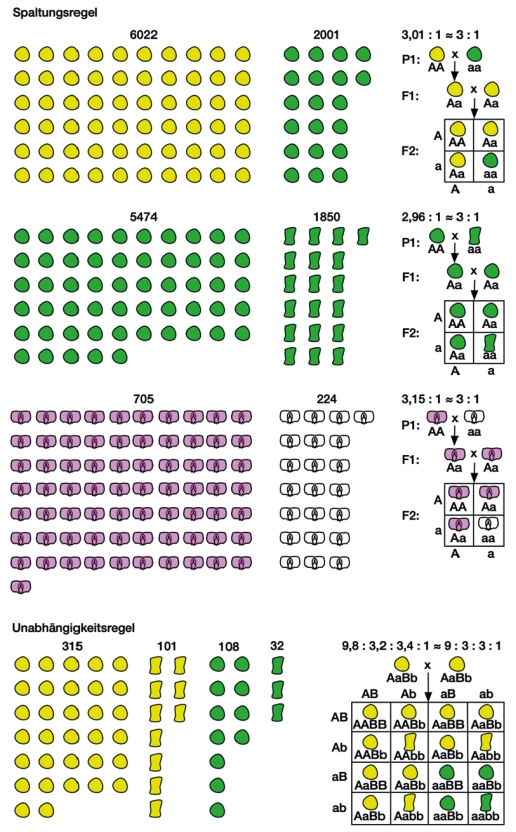
Abb. Anzahl der untersuchten Individuen und Spaltungsverhältnisse bei einigen der bekanntesten Experimente von Gregor Mendel (1866). Die Spaltungsregel konnte Mendel an Pflanzen mit gelben und grünen (oben), runden und eckigen Samen (Mitte) sowie rosa und weißblühenden Pflanzen (unten) bestätigen. Für die Unabhängigkeitsregel wählte er Samenfarbe (gelb und grün) sowie Samenform (rund und eckig). Oberhalb der Samen und Blüten stehen die Zahlenverhältnisse aus Mendels Versuchen, oberhalb der Kreuzungstabellen sind die beobachteten und die theoretischen Spaltungsverhältnisse dargestellt.
Mendel generalisierte seine Ergebnisse mit dem mathematischen Modell A + 2Aa + a (Mendel, 1866; Teicher, 2014). Heute würden wir mit der Erkenntnis, dass es sich bei A und a um zwei Allele desselben Gens handelt, die Schreibweise AA + 2Aa + aa wählen. Unabhängig davon beschrieb er damit, dass die Nachkommen der Kreuzung A × a, also Aa, bei Selbstung „die Neigung besitzen, zu den Stammeltern zurückzukehren" (Mendel, 1866). Die Beobachtung war nicht neu: Mendel schrieb, dass die „von Gärtner, Kölreuter und anderen gemachte Wahrnehmung" durch seine Versuche bestätigt wurde. Im Gegensatz zu seinen Vorgängern beließ Mendel es aber nicht bei einer „Wahrnehmung". Er postulierte, dass in jeder Selbstungsgeneration (n) das Verhältnis A:Aa:a = 2n-1:2:2n-1 ist. Das heißt, aus dem Verhältnis 1:2:1 in der ersten Filialgeneration der Hybride Aa wird das Verhältnis 1023:2:1023 in der zehnten Filialgeneration.
Vorhersagen dieser Art waren 1866 Standard in der Physik, aber neu in der Biologie, denn zur Zeit Mendels steckte die Anwendung von Mathematik in der Biologie noch in den Anfängen und wurde mehrheitlich eher skeptisch betrachtet (Keller, 2002; Deichmann, 2019). Zwar gab es bereits Bestrebungen Quantifizierung und Korrelation für den Bereich der Biogeografie und Pflanzenmorphologie anzuwenden, allerdings erlangte die Bedeutung der Statistik für die Biologie erst mit den Veröffentlichungen von Francis Galton (1884) und Karl Pearson (1900) ihren Durchbruch. Mendel war in dem Punkt den meisten Biologen seiner Zeit also einige Jahre voraus. Der Chi2-Test zum Beispiel, mit dem bis heute eine signifikante Abweichung von erwarteten Spaltungsverhältnissen statistisch abgesichert wird, wurde erst 34 Jahre nach Mendels Veröffentlichung von Pearson (1900) publiziert.
Mendel war kein etabliertes Mitglied der damaligen Wissenschaftsgemeinde und konnte seine experimentellen Ergebnisse und bahnbrechende Theorie zunächst nur im Rahmen von zwei öffentlichen Vorträgen in Brno vorstellen. Zwar verschickte er seine 1866 erschienene Publikation in einem Sonderdruck von 40 Exemplaren an verschiedene europäische Wissenschaftler, doch blieb dies weitestgehend ohne Resonanz (Orel, 1971). Bis 1900 wurde der Artikel von Gregor Mendel kaum beachtet und wahrscheinlich nur zweimal zitiert, ohne dass die Bedeutung seiner Arbeit dabei erkannt wurde (Zirkle, 1951).
Obwohl bereits vor Mendel mehrere Wissenschaftler Kreuzungsexperimente mit dem Fokus auf einzelne Merkmale durchführten, war seine Herangehensweise im Vergleich zu den meisten seiner Zeitgenossen in der Biologie eher unorthodox. Es war verbreitet, Spezies und Kreuzungen in ihrer Gesamtheit zu beschreiben, statt sich auf die Vererbung spezifischer Merkmale zu konzentrieren (Gasking, 1959). Ein weiteres Problem für die zunächst fehlende Beachtung der Theorie Mendels kann Darwins 1859 erschienenes Buch “On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life” gewesen sein. Das Buch führte zu großer Aufmerksamkeit und zu Kontroversen innerhalb der Wissenschaftsgemeinde und legte den Fokus auf die Beschreibung der natürlichen Selektion, die zunächst ohne gezielte Experimente zum Nachweis von Hypothesen auszukommen schien (Stubbe, 1963). Ohne komplett überzeugt zu sein, vertrat Darwin zudem die damals populäre blending inheritance-Hypthese, nach der sich die Merkmale beider Eltern in den Nachkommen mischen (Vorzimmer, 1963). Mendel selbst legte mit seinen Erbsenversuchen den Grundstein für die bis heute gültige particulate inheritance-Theorie, nach der sich die Merkmale beider Eltern nicht mischen, sondern neu kombinieren, (Mendel, 1866; Poczai & Santiago-Blay 2021). Dass Mendels Theorie die Evolutionstheorie Darwins stützte, wurde erst ab 1900 z. B. von Erich Tschermak (1900a) erkannt.
Zwar wies Rudolf Jacob Camerer schon 1694 anhand von Kreuzungsexperimenten die sexuelle Vererbung bei Pflanzen nach, allerdings konnte erst der Italiener Amici (1846) am Beispiel von Orchideen das Eindringen des Pollenschlauchs in den Embryosack und die Entwicklung des Embryos aus einem bereits dort vorhandenen ‚Bläschen‘, der Eizelle beobachten. Es sollte noch bis zum Jahr 1884 dauern, bis Eduard Strasburger die tatsächliche Verschmelzung des Spermazellkerns mit der Eizelle nachwies (Strasburger, 1884), aber spätestens mit Amici (1846) war Matthias Jacob Schleidens Theorie des Embryosacks als „Amme“ bereits vor Mendels Versuchen widerlegt (Lorenzano, 2011). Ausgerechnet Eduard Fenzl, Botaniker an der Universität Wien, war weiterhin ein Anhänger der Ammentheorie und ließ Mendel 1856 zum zweiten Mal durch die Lehramtsprüfung fallen (Lorenzano, 2011).
Bis zur „Wiederentdeckung“ der Vererbungsregeln durch Carl Correns (1900), Hugo de Vries (1900) und Erich Tschermak (1900a) sollten 34 Jahre vergehen. Während de Vries seine Gesetze zur Vererbung von Merkmalen zuerst publizierte, erkannte Correns, der zu ähnlichen Ergebnissen wie de Vries gekommen war, Mendel als wahren Erstautor der Vererbungsregeln an. Angeblich geschah das, nachdem er einen Sonderdruck von de Vries Forschungsarbeiten erhalten hatte (Wolpert, 2004). Laut Müller-Wille (2018) schrieb de Vries Mendel nur zu, die Vererbungsregeln für einen speziellen Fall dargestellt zu haben und nahm für sich selbst in Anspruch, die generelle Gültigkeit der Vererbungsregeln postuliert zu haben. Im Gegensatz dazu soll Correns in seinem in derselben Zeitschrift publizierten Artikel sarkastisch bemerkt haben, ihm käme es nicht auf die Erstveröffentlichung einer „Nachentdeckung“ an (Müller-Wille, 2018). Tschermak schreibt in der Nachschrift einer gekürzten Fassung seines Artikels (Tschermak, 1900b), ihn freue die "gleichzeitige Entdeckung Mendel's durch Correns, de Vries und mich". Er wird in der Literatur allerdings nicht durchgehend als "Wiederentdecker" der Mendelschen Regeln, gesehen (Simunek et al., 2012). Zweifelsfrei besonders bedeutend sind seine Arbeiten für die Entwicklung der Pflanzenzüchtung. Tschermak begründete 1909 an der Universität für Bodenkultur in Wien den ersten Lehrstuhl für Pflanzenzüchtung in Europa (Riedl-Dorn, 2016). Weitere internationale Aufmerksamkeit erhielten die Arbeiten von Gregor Mendel nachdem Wiliam Bateson im Jahr 1901 die Übersetzung des Artikels für die Royal Horticultural Society of London organisiert hatte (Fairbanks & Abbott, 2016).
Einigen Statistikern kamen die von Mendel erhobenen Spaltungsverhältnisse allerdings zu gut vor, um wahr zu sein. Bereits ein Jahr nach der englischsprachigen Veröffentlichung 1901 schrieb der Biometriker Raphael Weldon dem Mathematiker Karl Pearson, bezugnehmend darauf, dass die Beobachtungen des Augustinerabtes Gregor Mendel seinen postulierten theoretischen Spaltungsverhältnissen fast durchweg sehr nahekamen: „Remembering his shaven crown I cannot help wondering if they were not too good.“ Obwohl er etwas später schrieb, Mendel „cooked his figures, but (...) was substantially right“, hatte er weiterhin Bedenken gegenüber der Allgemeingültigkeit von Mendels Aussagen (Franklin, 2008), wobei er unter anderem Erich Tschermak-Seysenegg zitierte, der ebenfalls an dem Untersuchungsobjekt Erbse forschte, aber zumindest bei einigen der von Mendel untersuchten Merkmale weniger eindeutige Ergebnisse erzielte (Weldon, 1902; Tschermak, 1900a).
Weldon berechnete die Intervalle, innerhalb derer 50 % der Beobachtungen für die jeweils dominanten Eigenschaften in Abhängigkeit der Gesamtzahl der Observationen liegen müsste (0,75n ± 0,67449 × √n × 0,75 × 0,25). Bei den insgesamt sieben Versuchen mit unterschiedlichen Merkmalen, die Mendel (1866) zur Ableitung der Spaltungsregel durchführte, lagen seine Ergebnisse fünfmal innerhalb des Intervalls, einmal lag der Wert genau auf der Grenze und ein weiteres Mal lag er leicht oberhalb der Intervallgrenze. Weldon (1902) schlussfolgerte, die Chancen stünden 16:1, dass im Falle einer Wiederholung der Versuche schlechtere Ergebnisse erzielt würden. Mit seinen folgenden Versuchen hatte Mendel (1866) bestätigt, dass ein Drittel der Genotypen, die in der F2 das dominante Merkmal aufwies, bereits reinerbig sein muss. Basierend auf diesen Beobachtungen, berechnete Weldon (1902), dass die Werte von Mendel (1866) nur dreimal innerhalb des Intervalls lagen, einmal genau auf der Grenze und dreimal außerhalb der Grenze für den wahrscheinlichen Fehler. Er blieb skeptisch und bemerkte, Mendel habe nie Daten erhoben, deren Abweichungen von den theoretischen Spaltungsverhältnissen zweimal so groß wie der wahrscheinliche Fehler seien. Bezugnehmend auf die Versuche, in denen Mendel drei Merkmale untersuchte, kam Weldon (1902) zu dem Schluss, in 95 von 100 Fällen müssten bei wiederholten Versuchen schlechtere Ergebnisse erzielt werden. Bezugnehmend auf die zunehmende Popularität von Mendels Arbeiten schrieb Weldon (1902): „Some writers have been led to overlook the wonderfully consistent way in which Mendel’s results agree with his theory“.
Der später bekannt gewordene Statistiker und Genetiker Ronald A. Fisher soll 1911 im Alter von 21 Jahren ebenfalls berechnet haben, dass die Chancen bei einer Wiederholung der Experimente 16:1 gegen Mendel stünden, nochmals so gute Ergebnisse zu erzielen (Franklin, 2008). Er schrieb laut Franklin (2008): „It may just have been luck; or it may be that the worthy German abbot, in his ignorance of probable error, unconsciously placed doubtful plants on the side which favoured his hypothesis“. Als glühender Verfechter der Arbeiten von Gregor Mendel veröffentlichte Bateson zur selben Zeit gleich mehrere Bücher, die wiederum die Übersetzungen der Texte von Mendel, aber auch umfassende Abhandlungen zur Wiederentdeckung der Mendelschen Regeln und zur Allgemeingültigkeit der Vererbung von Merkmalen enthielten (Bateson, 1902; Bateson, 1913).
In einem 1936 publizierten Artikel kam Fisher zu dem Schluss, dass die meisten Daten Mendels verfälscht sein müssten, weil sie der Theorie zu gut entsprächen und größere Abweichungen zu erwarten wären. Bei seinen Untersuchungen der zweiten „Generation der Hybriden“ wählte Mendel (1866) jeweils 100 Pflanzen, die das dominante Merkmal zeigten und selbstete diese. Von jeder dieser Pflanzen nahm er zehn Samen, um die Spaltungsregel zu überprüfen. Insgesamt untersuchte er fünf Merkmale. Nur bei der Farbe der Hülsen stellte er eine größere Abweichung von der 2:1 Spaltungsregel fest: 60 der 100 ausgewählten Pflanzen bekamen sowohl Nachkommen mit gelben als auch solche mit grünen Hülsen. Sie waren demnach heterozygot. 40 bekamen ausschließlich Nachkommen mit grünen Hülsen und konnten als homozygot klassifiziert werden. Bei einer Wiederholung des Versuchs kam er zu einem Spaltungsverhältnis von 65:35, welches 2:1 wieder sehr nahekommt. Nimmt man jedoch die Nachkommen aller untersuchten 600 Pflanzen zusammen, waren 399 heterozygot und 201 homozygot für das untersuchte Merkmal. Die 2:1 Spaltungsregel war damit nahezu perfekt erklärt und könnte rein zufällig so gut abgebildet worden sein. Fisher (1936) ging allerdings davon aus, dass 5,63 % der heterozygoten Eltern des Selbstungsexperiments fälschlicherweise als homozygot klassifiziert werden müssten. Demzufolge sei das zu erwartende Spaltungsverhältnis nicht 2:1 sondern 1,7:1. Fisher (1936) schrieb, eine so glückliche Abweichung von den von ihm errechneten Spaltungsverhältnissen hin zu denen, die Mendel selbst postulierte sei nur in einem von 29 Experimenten zu erwarten.
Hartl & Fairbanks (2004) halten die Annahme Fishers allerdings für falsch, sobald ein kleiner Prozentsatz an heterozygoten Genotypen mit Merkmalen des rezessiven Phänotyps auftritt. Das würde für unvollständige Dominanzen zutreffen, die Mendel allerdings nicht beobachtet hat. Weiterhin argumentieren Sie, dass das Spaltungsverhältnis für die Hülsenfarbe dem erwarteten Verhältnis von Fisher sehr nahekommt, wenn man beide Versuche (Original und Wiederholung) zusammenfasst. Darüber hinaus gehen sie davon aus, dass Mendel als begnadeter Gärtner sicherlich mehr Pflanzen ausgesät hat, als er für seine Untersuchungen benötigte, um auf exakt zehn Nachkommen von den o.g. 100 Pflanzen zu kommen. Zudem sind einige der von Mendel untersuchten Merkmale aufgrund pleiotroper Geneffekte schon im Sämlingsstadium zu erkennen, sodass die Autoren hier von einer eher unbewussten Vorauswahl ausgehen. Weil keines der Ergebnisse von Mendel (1866) signifikant von dem erwarteten Spaltungsverhältnissen von 1,7:1 von Fisher (1936) abweicht, sehen Hartl und Fairbanks (2004) keinen weiteren Grund zur Beschuldigung Mendels, seine Daten bewusst gefälscht zu haben. Sind die Verdächtigungen also grundlos, wie Radick (2015) konstatierte? Es bleibt die Frage, warum außer 60:40 = 1,5:1 alle Abweichungen von dem von Fisher berechneten Spaltungsverhälnis von 1,7:1, welches die fälschlicherweise als homozygot klassifizierten heterozygoten Genotypen mit einbezieht, in dieselbe Richtung gehen. Sie sind also größer als 1,7:1 und nahe um das von Mendel postulierte Spaltungsverhältnis von 2:1 gestreut.
Für die in 1863 durchgeführten zwei- und dreifaktoriellen Experimente sowie für weitere Tests desselben Jahres gibt Fisher die erwarteten und beobachteten Abweichungen als Chi2-Werte an. Pearson (1900) beschrieb den Chi2-Test, der z. B. durch Harris (1912) propagiert wurde, um signifikante Abweichungen von Spaltungsverhältnissen nach den Mendelschen Regeln zu verifizieren. Fisher (1936) kommt zu dem Schluss, dass die Diskrepanz hochsignifikant ist und nur einmal in 2.000 Experimenten vorkommen könne. Alle Experimente zusammengefasst, kommt Fisher (1936) auf eine Wahrscheinlichkeit von 0,99993, mit der die erwarteten Abweichungen größer sind als die beobachteten. Das heißt, in nur sieben von 100.000 Fällen sind ebenso gute oder bessere Ergebnisse zu erwarten als die von Gregor Mendel. Nach der Durchführung von Monte-Carlo-Simulationen kommen Pires und Branco (2010) zu dem Schluss, dass die Ergebnisse von Fisher (1936) plausibel sind und gehen von zwei von 100.000 Fällen aus, in denen die Beobachtungen ebenso gut oder besser sein müssten als die von Mendel. Parallel analysierten sie die p-Werte anstelle der Chi2-Tests und berechneten, dass basierend darauf in vier von 1.000 Fällen einer Wiederholung der Experimente die Ergebnisse gleich gut oder besser zu den Erwartungswerten passen, als die von Mendel (1866). Die Conclusio von Pires & Branco (2010) ist, dass die Ergebnisse von Mendel „to good to be true“ seien, um alle Daten, die Mendel real in Versuchen erhoben hat, beinhalten zu können oder auf einer zufälligen Selektion der Datengrundlage zu basieren. Aufgrund seiner eigenen Äußerungen zu wiederholten Versuchen und der gelegentlichen Darstellung von extremen Abweichungen von der Regel, gehen Pires & Branco (2010) von einem „hidden bias“ aus, der zu den extrem positiven Ergebnissen von Mendel (1866) geführt hat.
Ganz im Gegensatz dazu nennen Montgomery & Birkhead (2005) Mendel den „father of scientific misconduct“ und spekulieren – wie bereits vorher vermutet, er könne viele Experimente durchgeführt, aber nur die veröffentlicht haben, in denen die Spaltungsverhältnisse nahe an seinen Erwartungen lagen. Zumindest in einem Fall benennt Mendel (1866) – wie oben beschreiben – konkret die Wiederholung eines Experimentes, weil die Ergebnisse zunächst nicht zu seinen Erwartungen passten. Bei Kreuzungsversuchen innerhalb der Gattung Phaseolus gibt Mendel nur an, „ein ganz übereinstimmendes Resultat“ beobachtet zu haben, ohne dieses im Detail zu dokumentieren. Als weitere Möglichkeit ziehen Montgomery & Birkhead (2005) in Betracht, Mendel könne aufgehört haben zu zählen, sobald die Zahlenverhältnisse seinen Erwartungen nahekamen, obwohl er selbst schrieb, nicht so vorgegangen zu sein. Auch könnten Pflanzen aussortiert worden sein, die zu sehr von den Vererbungsregeln abweichen. Bereits Fisher (1936) spekulierte, ein anonymer Assistent, der zu gut gewusst habe, was Mendel erwarte, könne Pflanzen beiseite genommen haben. Mendel selbst bemerkt an verschiedenen Stellen, dass bei den Nachkommen einzelner Pflanzen durchaus große Abweichungen von theoretischen Spaltungsverhältnissen zu beobachten sind. Dass diese nicht in die Analysen eingegangen sein könnten, geht aus seiner Schrift von 1866 allerdings nicht hervor. Auch Montgomery & Birkhead (2005) schließen die unbewusste Verfälschung der Daten, indem Mendel nicht eindeutig den Merkmalsausprägungen zuzuordnende Exemplare so einordnete, dass die theoretischen Spaltungsverhälnisse wieder stimmten, nicht aus.
Sollte Mendel seine Versuche unbewusst voreingenommen durchgeführt haben, hätte er schon vorab wissen müssen, welche Ergebnisse bzw. Spaltungsverhältnisse zu erwarten sind (Pilpel, 2007). Mit anderen Worten: Er hat seine Theorie über die Vererbungslehre nicht von den Ergebnissen abgeleitet, sondern die Ergebnisse – bewusst oder unbewusst – seiner Theorie angepasst. In der Wissenschaft ist es eher gewöhnlich, dass die Hypothese vor Beginn der Untersuchungen steht. Dass die objektive Herangehensweise oder der Versuch, die eigene Hypothese zu widerlegen, der korrekteste aber gleichzeitig emotional schwierigste Weg ist, dürfte jedem Wissenschaftler bewusst sein. Genau deshalb führen Pires & Branco (2010) die Notwendigkeit von Doppelblindstudien in einigen wissenschaftlichen Disziplinen an. Mit anderen Worten: Wenn Mendel wusste, dass ein weiterer runder Same zu erwarten war, hat er einen nicht ganz runden Samen eventuell als rund klassifiziert, der von einem komplett objektiven Beobachter als kantig bezeichnet oder als undefinierbar aussortiert worden wäre.
Ähnliches vermutet Weeden (2016), der davon ausgeht, Mendel habe ein brillantes Modell entwickelt und war deshalb versucht, dieses einem wenig enthusiastischen Publikum zu vermitteln. Weeden verglich die Chi2-Werte, die sich aus Mendels Daten ergeben mit denen, die später an denselben Merkmalen bei Erbsen beobachtet wurden. Er geht davon aus, dass zumindest bei einigen der untersuchten Merkmale deutlich stärkere Abweichungen von den theoretischen Spaltungsverhältnissen zu erwarten sind. Dabei beruft Weeden sich auf seine Metastudie, in der die Ergebnisse von 45 Publikationen eingingen (Tabelle). Zwar liegen die Ergebnisse von Gregor Mendel innerhalb der Grenzen der von Weeden zitierten Studien, allerdings ist auffällig, dass sie durchweg sehr gute bzw. den theoretischen Spaltungsverhältnissen von 1:1, 2:1 oder 3:1 nahekommende Ergebnisse zeigen. Darüber hinaus stellt sich die Frage, warum Mendel keine Kopplung von Merkmalen beobachtete, obwohl die Gene, die Wuchshöhe und Hülsenform kontrollieren, nur 5 cM entfernt auf demselben Chromosom liegen (Weeden, 2016). Dem Autor zufolge hätte Mendel diese Beobachtung machen müssen, die in seinem Artikel von 1866 unerwähnt bleibt. Allerdings berichtet Mendel von der Co-Segregation beider Merkmale in seinen Briefen an Carl Nägeli, die von Correns (1905) veröffentlicht wurden. Weeden schlussfolgert, wir sollten anerkennen, dass es keine unvoreingenommenen Beobachter gibt und dass Wissenschaft nur als Gemeinschaft selbstkorrigierend funktioniert.
Tabelle. Anzahl der von Mendel und anderen Autoren durchgeführten Versuche zu den Spaltungsverhältnissen verschiedener Merkmale bei der Erbse sowie durchschnittliche, minimale und maximale Chi2- und p-Werte. Bei der Tabelle handelt es sich um eine Zusammenfassung der Metastudie von Weeden (2016). Die 45 Publikationen, auf der die Studie beruht, sind im Originalartikel von Weeden (2016) zitiert. Ein Chi2-Wert > 3,84 bei einer Anzahl von Freiheitsgraden = 1 zeigt eine signifikante Abweichung von den theoretischen Spaltungsverhältnis 3:1 und 1:1. Dasselbe gilt für kleine p-Werte.
Merkmal | Mendel | andere Autoren (nach Weeden 2016) | ||||
Anzahl der Versuche | Chi2 | p | Anzahl der Versuche | Chi2 | p | |
Samenfarbe | 3 | 0.27 (0.10-0.39) | 0.62 (0.53-0.76) | 30 | 1.18 (0.00-5.00) | 0.44 (0.02-1.00) |
Wuchshöhe | 2 | 0.50 (0.39-0.61) | 0.49 (0.44-0.53) | 24 | 0.60 (0.00-2.11) | 0.57 (0.15-1.00) |
Hülsenfarbe | 1 | 0.45 (0.45-0.45) | 0.50 (0.50-0.50) | 33 | 1.50 (0.00-6.73) | 0.44 (0.01-1.00) |
Samenform | 8 | 0.14 (0.00-0.35) | 0.76 (0.56-1.00) | 28 | 2.18 (0.00-13.7) | 0.40 (0.00-0.98) |
Farbe der Kotyledonen | 8 | 0.31 (0.00-0.80) | 0.66 (0.37-1.00) | 15 | 1.60 (0.03-13.6) | 0.50 (0.02-0.86) |
Sitz der Blüten | 1 | 0.35 (0.35-0.35) | 0.55 (0.55-0.55) | 6 | 5.01 (0.00-7.68) | 0.33 (0.00-0.98) |
Hülsenform | 1 | 0.06 (0.06-0.06) | 0.80 (0.80-0.80) | 6 | 2.85 (0.20-11.4) | 0.35 (0.00-0.66) |
Letztlich bleiben also Zweifel an der korrekten Zählweise Mendels (1866) zur Erhebung seiner Daten bestehen. Ob er nun weitere Experimente durchgeführt und nur seine besten Ergebnisse veröffentlicht hat, ob einzelne Pflanzen durch ihn oder einen an keiner Stelle erwähnten Assistenten aussortiert wurden oder ob allein die fehlende Unvoreingenommenheit und Objektivität zu den vielen nahezu perfekten Ergebnissen geführt hat, wird wohl ungewiss bleiben und zu weiteren Spekulationen führen. Zu bedenken ist, dass Gregor Mendel im Gegensatz z. B. zu Charles Darwin kein bekannter Wissenschaftler war sondern – zum Zeitpunkt der Erhebung seiner Daten – ein einfacher Mönch, der seine Lehramtsprüfung nicht bestanden hatte. Trotzdem war seine quantitative Datenerfassung eher neu in seinem Forschungsgebiet und im Gegensatz zu seinen Vorgängern erkannte er die Gesetzmäßigkeiten bei der Vererbung von Merkmalen. Die von Mendel (1866) entdeckten theoretischen Spaltungsverhältnisse bleiben unangefochten und weisen gerade vor dem Hintergrund der Möglichkeiten, die Mendel für seine Forschungsaktivitäten hatte, auf seine Genialität hin. Mendel war der erste, der die Gesetzmäßigkeiten der Vererbung eindeutig beschreiben konnte und er hat damit die Grundlagen sowohl für die Genetik als Wissenschaft als auch für die moderne Pflanzenzüchtung gelegt.
Die Autoren erklären, dass keine Interessenskonflikte vorliegen.
Amici, G.B., 1846: Sulla fecondazione delle orchidee. Giornale Botanico Italiano. 2, 237–248.
Bateson, W., 1902: Mendel’s principles of heredity. A Defence, with a Translation of Mendel's Original Papers on Hybridisation. Cambridge University Press.
Bateson, W., 1913: Mendel’s principbles of heredity. 1. Mendels law 2. Evolution 3. Heredity 4. Hybridization. Cambridge University Press.
Correns, C., 1900: G. Mendels Regel über das Verhalten der Nachkommenschaft der Rassenbastarde. Berichte der Deutschen Botanischen Gesellschaft 18, 158–169.
Correns, C., 1905: Gregor Mendels Briefe an Carl Nägeli, 1866–1873. Ein Nachtrag zu den veröffentlichten Bastardierungsversuchen Mendels. Teubner, Leipzig.
Deichmann, U., 2019: From Gregor Mendel to Eric Davidson: Mathematical models and basic principles in biology. Journal of Computational Biology 26, 637-652, DOI: 10.1089/cmb.2019.0087.
de Rabutin-Chantal, M., 0000: “Potager du roi”, URL: https://de.wikipedia.org/wiki/Potager_du_roi.
de Vries, H., 1900: Das Spaltungsgesetz der Bastarde. Berichte der Deutschen Botanischen Gesellschaft 18, 83-90.
Fairbanks, D.J., S. Abbott, 2016: Darwin’s influence on Mendel: Evidence from a new translation of Mendel’s paper. Genetics 204, 401-405, DOI: 10.1534/genetics.116.194613.
Fisher, R.A., 1936: Has Mendel’s work been rediscovered? Annals of Science 1, 115–137, DOI: 10.1080/00033793600200111.
Franklin, A., 2008: The Mendel-Fisher Controversy. In: Franklin, A., A.W.F. Edwards, D.J. Fairbanks, D.L. Hartl, T. Seidenfeld (eds): Ending the Mendel-Fisher controversy. University of Pittsburg Press.
Galton, F., 1884: Regression towards mediocrity in hereditary stature. The Journal of the Anthropological Institute of Great Britain and Ireland 15, 246-263.
Gärtner, C.F., 1849: Versuche und Beobachtungen über die Bastarderzeugung im Pflanzenreich. KF Hering & Comp. Stuttgart.
Gasking, E.B., 1959: Why was Mendel’s work ignored? Journal of the History of Ideas 20, 60–84, DOI: 10.2307/2707967.
Goss, J., 1824: On the variation in the colour of peas, occasioned by cross-impregnation. Transactions of the Horticultural Society of London 5, 234-235.
Hartl, D.L., D.J. Fairbanks, 2004: Mud sticks: on the alleged falsification of Mendel’s data. Genetics 175 (3), 975-979, DOI: 10.1093/genetics/175.3.975.
Harris, J.A., 1912: A simple test of the goodness of fit of Mendelian ratios. The American Naturalist 46, 741-745.
Hedrick, U.P., F.H. Hall, L.R. Hawthorn, L. Berger, 1928: Peas of New York. Part 1 Vol. 1 of Vegetables of New York. Report of the New York State Agricultural Experiment Station. Albany JB Lion Company Printers.
Jefferson, T., 1766-1824: Garden Book. Original manuscript from the Coolidge Collection of Thomas Jefferson Manuscripts, Massachusetts Historical Society.
Keller, E.F., 2002: Making sense of life: Explaining biological development with models, metaphors, and machines. Harvard University Press.
Knight, T.A., 1799: Experiments on the fecundation of vegetables. Philosophical Transactions of the Royal Society of London 89, 195-204.
Knight, T.A., 1824: Some remarks on the supposed influence of the pollen in cross breeding. Transactions of the Horticultural Society of London 5, 377-380.
Kreplak, J., M.A. Madoui, J. Burstin, 2019: A reference genome for pea provides insight into legume genome evolution. Nature Genetics 51, 1411-1422, DOI: 10.1038/s41588-019-0480-1.
Lorenzano, E., 2011: What would have happened if Darwin had known Mendel (or Mendel’s work)? History and Philosophy of the Life Sciences 33, 3–49.
Mendel, G., 1866: Versuche über Pflanzen-Hybriden. Verhandlungen des Naturforschenden Vereins in Brünn 4, 3-47.
Miko, I., 2008: Gregor Mendel and the principles of inheritance. Nature Education 1, 134.
Montgomery, B., T. Birkhead, 2005: A beginner’s guide to scientific misconduct. ISBE Newsletter 17, 16-21.
Müller-Wille, S., 2018: Gregor Mendel and the history of heredity. In: Dietrich, M., M. Borrello, O. Harman (eds.) Handbook of the Historiography of Biology. 1-22, DOI: 10.1007/978-3-319-74456-8_8-1.
Orel, V., 1971: A reconstruction of Mendel's Pisum experiments and an attempt at an explanation of Mendel's way of presentation. Folia Mendeliana 6, 105–112.
Pearson, K., 1900: On the criterion that a given system of deviations from the probable in the case of a correlated system of variables is such that it can be reasonably supposed to have arisen from random sampling. Philosophical Magazine 50, 157-175.
Pilpel, A., 2007: Statistics is not enough: revisiting Ronald A. Fisher’s critique (1936) of Mendel’s experimental results (1866). Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 38, 618-626.
Pires, A.M., J.A. Branco, 2010: A statistical model to explain the Mendel-Fisher controversy. Statistical Science 25, 545-565, DOI: 10.1214/10-STS342.
Poczai, P., J.A. Santiago-Blay, 2021: Principles and biological concepts of heredity before Mendel. Biology direct 16, 1-17, DOI: 10.1186/s13062-021-00308-4.
Radick, G., 2015: Beyond the „Mendel-Fisher controversy“. Science 350, 159-160, DOI: 10.1126/science.aab3846.
Riedl-Dorn, C., 2016: "Tschermak von Seysenegg, Erich" Neue Deutsche Biographie 26, 475-476.
Seton, A., 1824: On the variation in the colour of peas from cross-impregnation. Transactions of the Horticultural Society of London 5, 236.
Simunek, M., U. Hoßfeld, O. Breidbach, 2012: 'Further Development' of Mendel's legacy? Erich von Tschermak-Seysenegg in the context of Mendelian-biometry controversy, 1901-1906. Theory in Biosciences 131, 243-252, DOI: 10.1007/s12064-012-0158-z.
Stern, C., E.R. Sherwood, 1966: The origin of genetics: a Mendel source book. WH Freeman San Francisco.
Strasburger, E., 1884: Neue Untersuchungen über den Befruchtungsvorgang bei den Phanerogamen als Grundlage für eine Theorie der Zeugung. Fischer, Jena.
Stubbe, H., 1963: Kurze Geschichte der Genetik bis zur Wiederentdeckung der Vererbungsregeln Gregor Mendels. Fischer, Jena.
Teicher, A., 2014: Mendel’s use of mathematical modelling: ratios, predictions and the appeal to tradition. History and Philosophy of the Life Sciences 36, 187-208, DOI: 10.1007/s40656-014-0019-9.
Townsend, B., 1725: The complete seedsman: Shewing the best and easiest method for raising and cultivating every sort of seed. London.
Tracy, W.W., 1903: American varieties of vegetables for the years 1901 and 1902. U.S. Department of Agriculture. Government Printing Office. New York.
Tschermak, E., 1900a: Über künstliche Kreuzung bei Pisum sativum. Berichte der Deutschen Botanischen Gesellschaft 18, 232-39 (gekürzte Fassung).
Tschermak, E., 1900b: Über künstliche Kreuzung bei Pisum sativum. Zeitschrift für die landwirtschaftlichen Versuchswesen in Österreich 3, 465-555.
Vorzimmer, P., 1963: Charles Darwin and blending inheritance. Isis 54, 371–390.
Weeden, N.F., 2016: Are Mendel’s data reliable? The perspective of a pea geneticist. Journal of Heredity 107, 635-646, DOI: 10.1093/jhered/esw058.
Weldon, W.F.R., 1902: Mendel's laws of alternative inheritance in peas. Biometrica 1, 228-254.
Wolpert, L., 2004: Unglaubliche Wissenschaft. Eichborn Verlag, Frankfurt am Main.
Woolridge, J., 1677: Systema horticulturae or the Art of Gardening. W. Freeman. London.
Zirkle, C., 1951: Gregor Mendel and his precursors. Isis: A Journal of the History of Science Society 42, 97-104.

 Suchen
Suchen