
Perspectives on long-term bee vitality monitoring
Perspektiven für ein Langzeit-Bienenvitalitätsmonitoring
Journal für Kulturpflanzen, 72 (5). S. 123–131, 2020, ISSN 1867-0911, DOI: 10.5073/JfK.2020.05.02, Verlag Eugen Ulmer KG, Stuttgart
 This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/deed.en).
This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/deed.en).Bees are essential pollinators and their protection is relevant to secure biodiversity and agricultural production. MonViA-project members and partners collaborate in monitoring projects to develop effective policies to support biodiversity in Germany. In the current case-study, the impact of climate on honey bee population performance was assessed. We modeled year-to-year Central-European honey yield changes and found + 1°C temperature change to stimulate annual honey yield by + 0.9 kg per colony, and + 100 mm precipitation to reduce honey yields – 0.4 kg. In regard to different climate change scenarios for Germany, our modelling suggests a potential + 0.4 to + 0.8 kg honey yield gain per colony in 2050, as compared to 2020. We conclude that the German honey bee population may benefit by rising temperatures. We discuss how bee performance is linked to weather and how our analysis would be strengthened by including more data, with a higher temporal and spatial resolution, i.e., intra-annually and -nationally. Pollinator trend monitoring should be extended with analyses that include e.g., extreme weather conditions, disease loads, availability of floral resource, beekeeping practice, land use and landscape structure.
Key words: Apis mellifera, honey bees, wild bees, pollinator populations, temperature, precipitation, climate, impact assessment
Bienen sind essentielle Bestäuber und daher ist ihr Schutz von zentraler Bedeutung für die Sicherung der biologischen Vielfalt und der landwirtschaftlichen Produktion. Innerhalb des Projekts MonViA werden viele Partner zusammenarbeiten, um wirksame Strategien zur Förderung der biologischen Vielfalt zu entwickeln. Wir präsentieren eine Langzeit-Fallstudie wie sich Klima auf die Leistungsfähigkeit von Honigbienenvölkern auswirkt. Veränderungen des mitteleuropäischen Honigertrags haben wir in Bezug auf die Änderung von Temperatur und Niederschlag modelliert. Eine + 1°C Temperaturänderung steigert den jährlichen Honigertrag um + 0,9 kg pro Volk, während + 100 mm Niederschlag den Ertrag um – 0,4 kg verringert. Basierend auf Klimawandelprognosen im Zeitraum 2020–2050, schätzen wir eine potenzielle Ertragssteigerung von + 0,4 bis + 0,8 kg Honig pro Volk. Wir schließen daraus, dass die Honigbienenpopulation in Deutschland von steigenden Temperaturen profitieren könnte. Weiterhin diskutieren wir, wie die Bienenleistung mit dem Wetter zusammenhängt und wie unsere Analysen durch die Einbeziehung weiterer Daten, mit einer höheren zeitlichen und räumlichen Auflösung, gestärkt werden könnten. Die Einflüsse extremer Wetterbedingungen, imkerlicher Praxis, Krankheitsbelastung, Verfügbarkeit von Nahrungsressourcen, Landnutzung und auch Landschaftsstrukturen sollten im Rahmen des Monitorings der Bienenvitalität miterfasst werden.
Stichwörter: Apis mellifera, Honigbienen, Wildbienen, Bestäuberpopulationen, Temperatur, Niederschlag, Klima, Folgenabschätzung
Honey bees and non-Apis wild bees provide the indispensable service of pollination, for both wild plants as well as crops. Protection of bees therefore is of utmost relevance to human nutrition, society and the environment. The need and urgency for protective measures is high considering the progressive loss of biodiversity, involving decline of vegetation, animals and interlinking ecosystem functions, such as bee-pollination (Kevan and Viana, 2018; IPBES, 2019; Diaz et al., 2019). Earth’s entomofauna shows steep declines in many regions (Biesmeijer et al., 2006; Hallmann et al., 2017; Bidau, 2018; Lister and Garcia, 2018; Powney et al., 2019; Forister et al., 2019; Seibold et al., 2019; Hallmann et al., 2020). However, a baseline with spatially inclusive and comprehensive data is missing, and much needed (Didham et al., 2020). To protect and improve vitality and diversity of bee populations on the long term, we need general trend monitoring to find robust indicators to develop effective measures, to support the performance and vitality of bee-pollinators.
Several factors are implicated in the decline of pollinator populations, including habitat loss (Seibold et al., 2019), landscape fragmentation, pesticide use (Woodcock et al., 2017), parasites and pathogens (Schroeder and Martin, 2012; Cornelissen et al., 2019), and climate change (Sánchez-Bayo and Wyckhuys, 2019) as main drivers concerning bees. In terms of floral resource continuity, especially habitat and climate are major drivers in provisioning bees with a balanced and high-quality nutrition (pollen and nectar), in particular at times of rearing offspring. Intensive agricultural production systems often cannot cover nutritional needs for pollinator populations – which may contribute to inadequate crop pollination (Thompson, 2001). The current trend of declining pollinator abundancies within agricultural landscapes may be countered by cross-linking habitats with near-natural vegetation and flowering strips or hedgerows (Schulte et al., 2017; Dolezal et al., 2019; Seibold et al., 2019).
With this study, we aim to provide perspective on long-term monitoring of honey bee vitality, regarding many interactions, e.g. between land use, climate and bee health. Populations are influenced by many factors that may change over time, thus trend monitoring using long-time data series will help to understand the driving factors to vital pollinator populations. Our broader research scope is framed within a nationwide monitoring of biodiversity in agricultural landscapes called MonViA (see Table 1). Project MonViA aims to investigate which policy measures have a positive influence on agricultural biodiversity, including bee population health and diversity. A large scale monitoring can provide a strong scientific basis for determining measures to promote and fund affirmative action, towards safeguarding appropriate habitats with sufficient resources for bees.
Table 1. MonViA project overview.
| National Monitoring of Biological Diversity in Agricultural Landscapes – a project on behalf of the German Federal Ministry of Food and Agriculture, involving the Federal Agency for Agriculture and Food, the Thünen-Institut and Julius Kühn-Institut. |
Why MonViA | Land area in Germany is 50% used for agriculture. According to many studies, agriculture is a key driver of biodiversity decline, and biodiversity changes affect the performance and stability of agricultural production. Key Question: Which measures need to be recommended by policy makers to promote biodiversity, e.g., of bee pollinators? |
What we do | The Institute for Bee Protection (Julius Kühn-Institut) works on five subprojects. 1) Monitoring of honey bees: Extensive data from various projects exist, e.g. on honey yields, bee health parameters, and population densities. We aim to combine and perform multivariate long-term trend analyses. 2) Monitoring of wild bees: We aim to facilitate the central collection of nationwide recording data on wild bees: from literature research, expert reports, and from collectors. 3) Hymenopterans in orchards and vineyards: A systematic study on management practice impacts on the biodiversity of wild bees and wasps. 4) Citizen Science: A public mobile phone application will collect site-specific data on honey bee and wild bee occurrences. 5) Digital platform: Current and historic data on wild and honey bees, as well as climate and land use, are to be brought together in a national database. The repository is used to improve insight into the effects of land use on distribution and vitality of bees, and to enable regional assessment of pollination services. |
How | We will cooperate with many German partners to encompass regional and national datasets on wild and honey bees from institutions, experts, farmers, beekeepers, and citizen scientists. |
Need | Long-term, continuous, comprehensive biodiversity data collection for Germany. Within MonViA, monitoring data are harmonized with other national and international monitoring programmes, to allow reciprocal support and advantage. |
The complexity of climate change is currently much in focus of biodiversity concerns, because not only average weather conditions are shifting, but also extremes such as heat and droughts are increasing (Wagner, 2020). Global warming may cause mismatched plant-pollinator interactions (Scaven and Rafferty, 2013; Gérard et al., 2020). When native ranges become inhabitable due to climate change, both plants and pollinators may need to shift their habitat ranges, if possible, in particular for highly adapted species. Some native bee species, such as bumble bees in North America and Europe, have been reported to be pushed toward more Northern ranges, or otherwise become extinct (Kerr et al 2015; Soroye et al., 2020). Alternatively, other bee species may profit from a warmer climate. For instance, the Western honey bee (Apis mellifera) is an African/Mediterranean species in its evolutionary origin (Whitfield et al., 2006), hence this bee potentially performs better if summers in Central-Europe become warmer and dryer.
In the current case-study, we explore to what extent climate affects honey bee colony productivity in Germany. Honey yield per colony is a good proxy to measure colony vitality, because it is based on colony health and the availability of forage in the surrounding habitat. We expect increasing temperature (T) to be beneficial to honey yields, for instance via stimulating bee activity, and by stimulating the growth of floral resources. Precipitation (P) is also essential to colony development, in regard of floral resource development. Notwithstanding, honey bee foraging may be impeded by long periods of rainfall, or by severe drought, thus precipitation may also be a restrictive variable to honey yields (He et al., 2016). The derivative question is to what extent the whole population is affected by climate change. To answer this, we model the relationship between environmental conditions and colony performance for Central Europe in general, and predict long-term change in colony performance due to climate change for Germany in particular.
Honey yield per colony – a proxy for colony performance – was analyzed with linear modeling. We describe how year-to-year honey yield is affected by average annual weather variables. The analysis was performed on relativized data (see below), independent of long-term changes in honey production (e.g., differences in beekeeping practice, economy, emerging pests, data collection, etc.).
Honey production and bee colony data (1961–2017) was taken from the FAO database (http://faostat.fao.org), which is based on data collected by the Deutscher Imkerbund e.V. (D.I.B.- Mitgliederstatistik). We calculated yearly honey yield (Y) data by dividing honey production (in tons) by population size (number of hives). Weather data, i.e., yearly average temperature and precipitation values for the period 1961–2017 were taken from the CEDA archive (http://archive.ceda.ac.uk; CRU CY 4.02). Temperature data (T) were in °C, and precipitation data (P) were in millimeters (see Fig. 1E; 1 mm = 1 L/m2), with precipitation effect size summarized into decimeters (see Fig. 2B; 1 dm). The weather impact analysis on honey yields was performed using data of six Central European countries (EU; n = 342). The countries were Germany and its neighbors (DE, FR, CH, AT, TCH, PO), excluding countries with insufficient data (DK, NL, BE, LU). We fused data of Czech Republic and Slovakia into former Czechoslovakia (TCH) to have a continuous data set for 1961–2017 (Moritz and Erler, 2016).
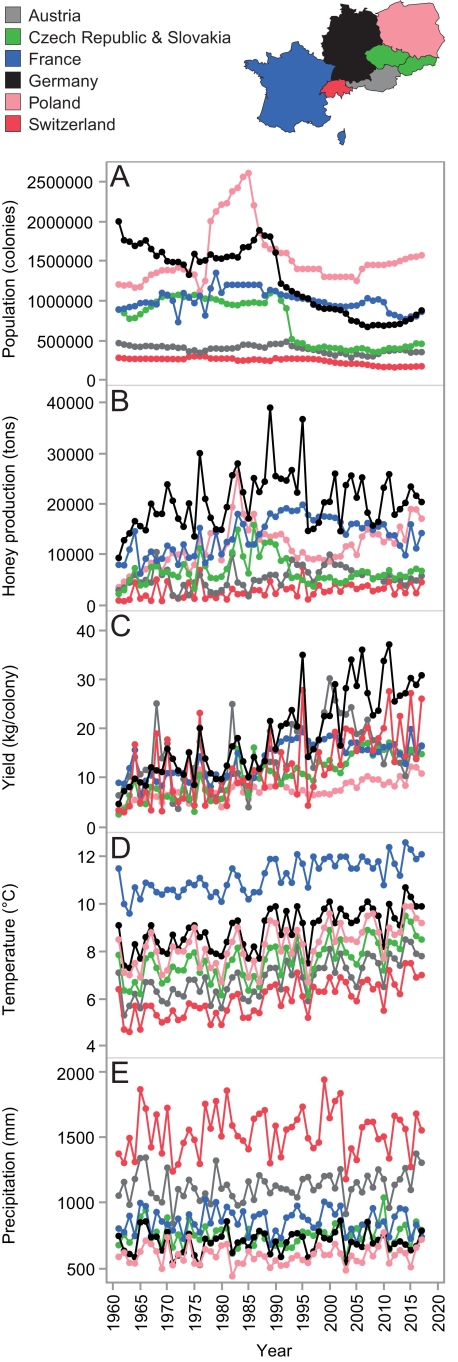
Fig. 1. Central-European honey bee and weather data. Shown are annual data on total honey bee populations (A), total honey productions (B), mean colony yields (C), mean temperatures (D), and mean precipitations (E). Lines connect dots of sequential yearly data, per country (see legend), for the period 1961–2017.
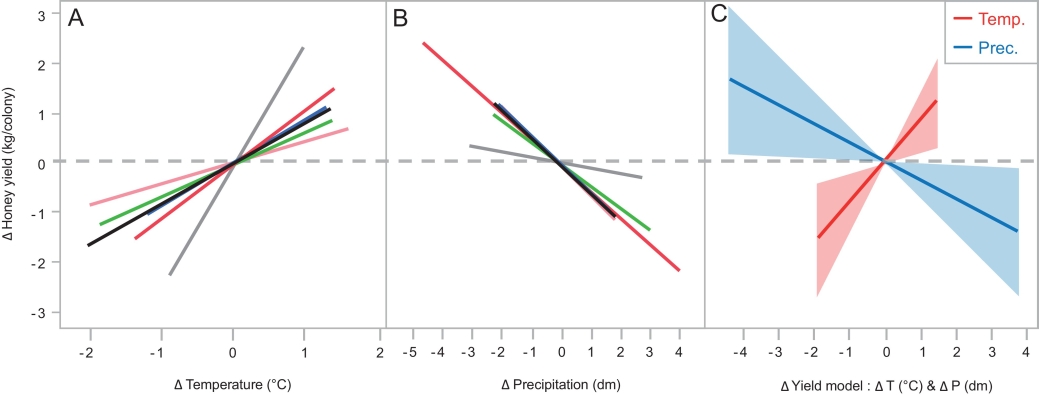
Fig. 2. Impact of annual weather on honey yields. Differences in honey yield ΔY are shown in relation to variance in yearly temperatures ΔT (A), and precipitationsΔP (B) in six Central-European countries (colors shown at Fig. 1). The lines with shaded 95%-CI intervals (C), indicate temperature (red) and precipitation (blue) deviation effects in our year-based honey yield model;ΔY = 0.87 × ΔT – 0.39 × ΔP.
Data analyses and modeling were performed using JMP Pro 15.1.0 software. For Germany, Temperature (T) and precipitation (P) data were analyzed over time by means of linear regression (1961–2017; n = 57 years). Linear Mixed Effects Models were used to assess if honey yields in Central Europe were relatively lower or higher (ΔY) according to years being relatively colder or warmer (ΔT) and/or wetter or dryer (ΔP). The data ΔY, ΔT and ΔP were calculated per country and per whole year, relative to a baseline average of ± 5 neighboring years, e.g., ΔY year x = Yyear x minus the decade average (Yyears x-5, x-4, x-3, x-2, x-1, x+1, x+2, x+3, x+4, x+5). This weather impact analysis (EU model) included ΔY as response variable, with the predictor variables ΔT and ΔP. We further included country identity as random factor (6 levels), and three interaction terms: ΔT × ΔP, ΔT × Country, ΔP × Country. Non-significant interactions (p > 0.05) were removed from the model. The model intercept was set to zero since all data values were relative and on average zero.
Climate change prediction for Germany are listed in Table 2. A best-, an intermediate-, and a worst-case greenhouse gas concentration scenario (Representative Concentration Pathway (RCP) 2.6, 4.5 and 8.5, respectively) were provided by Germany's National Meteorological Service (Deutscher Wetterdienst). We estimated yearly temperature and precipitation averages for years 2020 and 2050, and the change (Δ) over this period, by linear interpolations of baseline values for the period 1971–2000 (8.5°C and 773 mm) and the climate predictions for the period 2031–2060 (Table 2).
Table 2. Climate predictions for Germany. Temperature (T; in °C) and precipitation (P; in mm) predictions, given for a best-, intermediate-, and worst-case scenario (RCP 2.6, 4.5, and 8.5). We interpolated data between the periods 1971–2000 (baseline) and 2031–2060, to obtain estimates for years 2020 and 2050, and change (Δ) estimates for this period (in bold). Given values are medians (50th percentile) with a range indication in brackets [15th percentile – 85th percentile].
Climate Predictions | RCP 2.6 | RCP 4.5 | RCP 8.5 |
T period 2031–2060 | 9.7 [9.5 – 10.1] | 10.0 [9.6 – 10.4] | 10.5 [10.2 – 10.8] |
T interpolation year 2020 | 9.2 [9.1 – 9.4] | 9.4 [9.2 – 9.6] | 9.7 [9.5 – 9.8] |
T interpolation year 2050 | 9.8 [9.6 – 10.1] | 10.1 [9.7 – 10.6] | 10.7 [10.3 – 11.0] |
ΔT 2020 – 2050 | + 0.6°C | + 0.7°C | + 1.0°C |
P period 2031–2060 | 815 [775 – 842] | 822 [789 – 851] | 825 [775 – 857] |
P interpolation year 2020 | 797 [774 – 813] | 801 [782 – 818] | 803 [774 – 821] |
P interpolation year 2050 | 818 [775 – 847] | 825 [790 – 857] | 829 [775 – 863] |
ΔP 2020 – 2050 | + 20.9 mm | + 24.3 mm | + 25.8 mm |
The annual population size of honey bee colonies in Germany over the period 1961–2017 was on average 1.3 million hives ± 0.42 SD with an average of 21 thousand tons ± 5.5 SD honey produced per year (n = 57) (Fig. 1; A, B). The yearly honey yield per colony was on average 18.8 kg ± 8.6 SD. Honey yields in Germany are increasing, with + 0.44 kg annually (Fig. 1C; F1,55 = 144.5, p < 0.0001, R2 = 0.72, Formula: Yield = –853 + 0.438 × year). The absolute honey yield data showed annual autocorrelation (ACF = 0.71), i.e., the honey yield results of one year tended to be similar to proceeding and/or succeeding years.
Over the period 1961–2017 the yearly mean temperature and precipitation in Germany were 8.9°C ± 0.8 SD, and 699 mm ± 83 SD, respectively. The average temperature in Germany increased significantly, with 1.9°C (Fig. 1D; F1,55 = 46.2, p < 0.0001, R2 = 0.46, Formula: T = –58.9 + 0.034 × year). Precipitation did not significantly change (+23 mm) over the period 1961–2017 (Fig. 1E; F1,55 = 0.34, p = 0.55, R2 = 0.006, Formula: P = –80.7 + 0.39 × year).
Regarding the data of six Central-European countries, we found years with high temperature to stimulate honey yield (Fig. 2A; ΔT: F1,339.3 = 7.07, p = 0.008), while a high precipitation to reduce yield (Fig. 2B; ΔP: F1,323.7 = 4.69, p = 0.031). The interaction ΔT × ΔP showed that temperature and precipitation affected honey yields independently (F1,4.88 = 1.35, p = 0.30). The interaction ΔT × Country showed that yield stimulation by temperature is a valid effect for Central Europe in general, since the effect did not differ between particular countries (F1,319.2 = 0.86, p = 0.51). The interaction ΔP × Country result showed that yield decline by precipitation is also a valid effect for Central Europe in general with no difference between countries (F5,323.3 = 0.18, p = 0.97). The three non-significant interactions were removed from the model.
The Central-EU-wide model found + 1°C temperature change to stimulate honey yield with + 0.9 kg per colony, while + 1 dm precipitation change to reduce honey yield with – 0.4 kg per colony (Fig. 2C). A low mean R2 value of 0.08 (n = 6 countries) suggests a weak model fit, with 8% annual yield variation explained by the annual average temperature and precipitation data, and the remaining 92% being unexplained variation.
The EU weather model coefficients (Fig. 2C), combined with a 30 year climate prediction for Germany (Table 2), predict honey yield per colony to increase with + 0.4 kg, + 0.5 kg or + 0.8 kg, due to climate change, under best-, intermediate-, and worst-case green house gas concentration scenarios, respectively. With the approximate 0.9 million hives in Germany to date, these scenarios reflect a potential national production increase of 383 tons, 484 tons and 677 tons honey in 2050 as compared to 2020, due to climate change. These predictions come with the cautionary note that we ignored autocorrelation trends within temporal datasets, and that we based the predictions on linear relation assumptions.
Considering the high economic and natural value of pollination, it is important to maintain healthy bee populations to safeguard adequate pollination for crops and wild plants. However, bee pollinators globally are facing problems, with reduced biodiversity and declines in populations (Potts et al., 2010; Kevan and Viana, 2018). In a case study for Germany, we investigated long-term honey bee colony performance, with a focus on potential effects of climate change. Our model for Central Europe indicated a significant positive impact of higher annual temperature on honey yield, as well as a significant negative impact of annual precipitation on honey yield. Subsequently, changing climate variables (Table 2) might affect the German honey bee population positively, as our estimates predict an increase in honey production.
We suspect two underlying principles to drive effects on honey yield, as measure of honey bee performance. On the one hand, climate affects vegetation development, thus determines the availability and quality of floral resources for bees (Ogilvie et al., 2017). On the other hand, daily weather conditions can affect bee foraging behaviors directly, i.e., by temperature (Aublet et al., 2009; Comba, 1999), rain (He et al., 2016), but also potentially by wind (Comba, 1999; Hennessy et al., 2020). However, in absence of day-to-day environmental data, with parallel data on colony growth, our case-study may merely suggest the causation of climate impact on yearly colony performance (Fig. 2). Additional analyses per season may further differentiate optimal performance conditions for colonies, noting that warmth may stimulate honey yields (this study), yet it may also detriment colony survival during the subsequent winter (Switanek et al., 2017). We also note that the largest part in yield data variation remained unexplained. Thus, our aim in context of the MonViA-project is to combine a multitude of long-term data sets, with day-to-day data sets, to improve the impact assessments for bee performance.
In this study, we find that honey yields in several Central European countries respond similarly to temperature and precipitation impacts (Fig 2). Our results are specific to the central-EU region, i.e., as perhaps more rain may benefit colonies in dryer climates, but not in wetter climates. Similarly, very cold regions may benefit by a temperature increase, while very hot regions may not. We recommend and intend to extend this research further, with additional data of colder and hotter regions included, to estimate climatic ranges for optimal honey bee colony performance, and for assessing the consequences of seasonally dependent changes. Further, a non-linear approach may allow to help identify performance optima, for which we need more data, covering large spatio-temporal scales.
We aim to include vegetation data into performance analyses, to disentangle direct climatic effects on colonies, from indirect climatic impacts via the development and availability of floral resources (nectar and pollen). Regarding vegetation, we note that with 16.7 million hectares about half of the total area of Germany is under agricultural land use. Our bee performance assessments, in context of the MonViA project, will particularly consider agricultural landscapes. For land use and landscape structure are various long-term monitoring data already available (e.g., crops grown, production rates, phenology, intensive versus extensive management practices, use of agrochemicals such as pesticides and fertilizers, near-natural habitats, and heterogeneity and connectivity of landscapes). The combination of continuous datasets via beekeeping statistics, meteorological data, and plant-phenology data, should enable to separate direct and indirect impacts by weather on bees.
Within MonViA, we strive a 100% spatial coverage for Germany, with spatial and temporal extensions of datasets, by means of including historical data and international data, and combining academic data with federally collected statistics, and other available data, like from monitoring by remote sensing. As illustrated with our multi-national data approach (Fig. 2), international context is valuable to corroborate regional or national findings. International data can enrich analyses of data that are not nationally available. For example, colony performance in light of an ongoing epidemiological monitoring of multiple honey bee diseases in Switzerland (von Büren et al., 2019), might be transferable to the German situation.
Noteworthy is the opportunity to investigate the interaction between climate and bee pathogens. As disease pathogeneses can be linked to weather conditions, global warming may indirectly impact bee health and vitality. For example, the reproduction of Varroa destructor, a serious brood parasite of honey bee colonies is highly linked to climatic conditions during the winter (Nürnberger et al., 2019). Ascosphaera apis (chalkbrood) may manifest itself by larval mycosis when bee larvae get chilled (Puerta et al., 1994). This disease is common in social bees (Aronstein and Murray, 2010), but also in solitary bees (Torchio, 1992; James, 2005). Understudied is also the pathogenesis of bee viruses that plague colonies, and their potential interaction with climate (Di Prisco et al., 2011; Natsopoulou et al., 2017). It is possible that, especially at times of nutritional stress and/or under extreme climatic conditions, certain diseases may prosper and contribute to the demise of bee populations.
Managed honey bees undeniably provide essential pollination services, yet the high diversity of species and families of solitary bees deserve more credit and attention as regarding their importance for pollination in agro-ecosystems. As compared to managed honey bees, wild bees are relatively understudied and it is difficult to perform long-term trend monitoring regarding their population statuses. If data sets on honey bees and wild bees are combined, with inclusion of historical data, as well as with data from current monitoring projects, meta-level analyses opportunities are enabled. For example, regarding stability and vitality of parallel living bee populations in Germany, over time, a positive trend observed for one bee population may be shared by another, for instance in response to floral improvement via global warming. In contrast, negative interaction of concern are that certain bees may outcompete others via floral resource collection (Steffan-Dewenter and Tscharntke, 2000; Mallinger et al., 2017), and that pathogens can be transmitted between species (Dolezal et al., 2016; Graystock et al., 2015; Mallinger et al., 2017), meanwhile impacting populations of bees, and plants, differentially (Gérard et al., 2020).
To our perspective, many of the factors that drive honey bee colony productivity, may hold validity for the performance of wild bees too, e.g., regarding floral resources, pesticide exposure, diseases, etc., alongside broader societal effects like land-use change. To date, the majority of studies into pollinator declines focus on honey bees, and to a lesser extent on bumble bees. For Germany, but also for the world in general, there is a notable lack of information on the solitary bees, which make up the majority, over 85%, of the estimated 25,000–30,000 species of bees worldwide (Batra, 1984). The identification of wild bees is time-intensive and expertise is very scarce, resulting in datasets which tend to be regional and not open-source. However, in light of progressive ease of data-management in this age of digitization, we aim to direct effort to make expert data on wild bee populations available, to perform inter-regional trend monitoring (i.e., national and international). The multitude of investigations on individual, population, and ecosystem levels should enable a scientific data driven approach to support insect populations.
The use of an overarching data platform, as foreseen in project MonViA, envisions further integrations with pollinator relevant datasets. For honey bees, the integration of epidemiological data on bee diseases, pesticide damages, and beekeeper data collections, are valuable for the monitoring of long-term population health measures. In addition, spatio-temporal use of pesticide application data in agriculture may reveal the impact of detrimental agrochemical applications. Integration of land use data, such as remote sensing data on crops and landscape structures, may help explain inter-regional variation in bee performance.
With the current momentum of increasing computation power, and genetic data analyses capacities, also population genetics are valuable metrics to consider, for instance in regard of the stability and resilience of environmentally challenged populations (Matias et al., 2017; Theodorou et al., 2018; Theodorou et al., 2020). Mating frequency of honey bee queens reduce under more rainy conditions (El-Niweiri and Moritz, 2011), which may negatively affect queen performance (e.g., egg laying), and the lower degree of polyandry may limit colony performance (i.e., due to a low gene diversity and lower disease resistance). For honey bees in particular, different breeding lines are a very interesting variable to consider in regard to colony performance. Some strains may be climatically better adjusted, while others may perform relatively better under pressure of disease and parasites, e.g., Varroa destructor (Nürnberger et al., 2019).
Many citizen science initiatives represent great opportunities in support of bee population monitoring. Data collected by lay persons can support the analyses of scientific data collections (Brodschneider et al., 2019). Ecosystem monitoring efforts for wild bees can be supported via, e.g., species identification for biodiversity assessment. For honey bees, the involvement of beekeepers as citizen scientists is extremely valuable. Data on colony health can flow both from and toward beekeepers, noting that honey bee colony health depends on the extent to which beekeepers are informed on beekeeping practices (Jacques et al., 2017). Access to honey bee colony data is essential within the MonViA project, for instance, to allow making resilience estimates. Whereas small colonies are likely relatively vulnerable, large colonies may be able to buffer climatic impact and better bridge times of nutritional scarcity. Yet above all, honey bee colonies are managed, which involves essential information regarding beekeeping practices. The feeding of colonies, the treatment of parasites and diseases, the travel with colonies toward floral resources, are all beekeeping interventions, likely highly explanatory to colony growth, productivity and survival. Hence, an essential component in the analyses of honey bee colony performances is the consideration of beekeeping practices (Genersch et al., 2010).
With the MonViA project we aim to provide an internationally linkable data resource for agricultural landscapes and beyond, to monitor national long-term biodiversity trends, in support pollinators and safeguard biodiversity as a whole. Based on our modelling results, we conclude that the honey bee population in Germany could potentially benefit by rising Central-European temperatures. Albeit, population level data analyses based on yearly country averages (Fig. 1) are most certainly less powerful to assess environmental impacts on colony performance, as compared to within year colony level data. Notwithstanding, the combination of different research approaches can enable to bridge knowledge gaps, e.g., between short-term causality findings, and observed long-term population trends.
The presented study is part of the joint project ‘Farmland biodiversity Monitoring’ (MonViA) that has been funded by the Federal Ministry of Food and Agriculture. We are much obliged to Stephan Härtel and Robin F. A. Moritz for reviewing the manuscript before submission. Torsten Ellmann and Olaf Lück are kindly thanked for sharing Deutscher Imkerbund e.V. Mitgliederstatistik data. Thomas Leppelt from the Deutscher Wetterdienst (Abteilung Agrarmeteorologie) is kindly thanked for providing RCP scenario-based climate model predictions. We are grateful to Abdulrahim T. Alkassab, André Krahner and Jens Pistorius for their input and discussion.
The authors declare no conflicts of interest.
Aronstein, K.A., K.D. Murray, 2010: Chalkbrood disease in honey bees. Journal of Invertebrate Pathology 103, S20–S29, DOI: 10.1016/j.jip.2009.06.018.
Aublet, J.F., M. Festa-Bianchet, D. Bergero, B. Bassano, 2009: Temperature constraints on foraging behaviour of male Alpine ibex (Capra ibex) in summer. Oecologia, 159 (1), 237e247.
Batra, S.W.T., 1984: Solitary Bees. Scientific American 250, 120–127.
Bidau, C.J., 2018: Doomsday for insects? The alarming decline of insect populations around the world. Entomology, Ornithology and Herpetology 7, 1000e1130, DOI: 10.4172/2161-0983.1000e130.
Biesmeijer, J.C., S.P. Roberts, M. Reemer, R. Ohlemüller, M. Edwards, T. Peeters, A.P. Schaffers, S.G. Potts, R. Kleukers, C.D.O. Thomas, J.O. Settele, W.E. Kunin, 2006: Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 313 (5785), 351-354, DOI: 10.1126/science.1127863.
Brodschneider, R., K. Gratzer, E. Kalcher-Sommersguter, H. Heigl, W. Auer, R. Moosbeckhofer, K. Crailsheim, 2019: A citizen science supported study on seasonal diversity and monoflorality of pollen collected by honey bees in Austria. Scientific Reports 9, 16633, DOI: 10.1038/s41598-019-53016-5.
Clarke, D., D. Robert, 2018: Predictive modelling of honey bee foraging activity using local weather conditions. Apidologie 49, 386–396, DOI: 10.1007/s13592-018-0565-3.
Comba, L., 1999: Patch use by bumblebees (hymenoptera apidae): Temperature, wind, flower density and traplining. Ethology Ecology and Evolution, 11 (3), 243e264, DOI: 10.1080/08927014.1999.9522826.
Cornelissen, B., P. Neumann, O. Schweiger, 2019: Global warming promotes biological invasion of a honey bee pest. Global Change Biology 25 (11), 3642-3655, DOI: 10.1111/gcb.14791.
Didham, R.K., Y. Basset, C.M. Collins, S.R. Leather, N.A. Littlewood, M.H.M. Menz, J. Müller, L. Packer, M.E. Saunders, K. Schönrogge, A.J.A. Stewart, S.P. Yanoviak, C. Hassall, 2020: Interpreting insect declines: seven challenges and a way forward. Insect Conservation and Diversity 13, 103-114; DOI: 10.1111/icad.12408.
Di Prisco, G., X. Zhang, F. Pennacchio, E. Caprio, J. Li, J.D. Evans, G. DeGrandi-Hoffman, M. Hamilton, Y.P. Chen, 2011: Dynamics of Persistent and Acute Deformed Wing Virus Infections in Honey Bees, Apis mellifera. Viruses 3 (12), 2425-2441, DOI: 10.3390/v3122425.
Diaz, S., J. Settele, E.S. Brondízio, H.T. Ngo, J. Agard, A. Arneth, P. Balvanera, K.A. Brauman, S.H.M. Butchart, K.M.A Chan, L.A. Garibaldi, K. Ichii, J. Liu, S.M. Subramanian, G.F. Midgley, P. Miloslavich, Z. Molnár, D. Obura, A. Pfaff, S. Polasky, A. Purvis, J. Razzaque, B. Reyers, R.R. Chowdhury, Y.J. Shin, I. Visseren-Hamakers, K.J. Willis, C.N. Zayas, 2019: Pervasive human-driven decline of life on Earth points to the need for transformative change. Science, 366 (6471), eaax3100, DOI: 10.1126/science.aax3100.
Dolezal, A.G., S.D. Hendrix, N.A. Scavo, J. Carrillo-Tripp, M.A. Harris, M.J. Wheelock, M.E. O’Neal, A.L. Toth, 2016: Honey Bee Viruses in Wild Bees: Viral Prevalence, Loads, and Experimental Inoculation. PLOS ONE 11 (11), e0166190, DOI: 1371/journal.pone.0166190.
Dolezal, A.G., A.L. St. Clair, G. Zhang, A.L. Toth, M.E. O’Neal, 2019: Native habitat mitigates feast–famine conditions faced by honey bees in an agricultural landscape. Proceedings of the National Academy of Sciences, 116 (50), 25147-25155, DOI: 10.1073/pnas.1912801116.
El-Niweiri, M.A.A., R.F.A. Moritz, 2011: Mating in the rain? Climatic variance for polyandry in the honeybee (Apis mellifera jemenitica), Population Ecology 53, 421-427, DOI: 10.1007/s10144-011-0271-8.
Forister, M.L., E.M. Pelton, S.H. Black, 2019: Declines in insect abundance and diversity: we know enough to act now. Conservation Science and Practice 1, e80, DOI: 10.1111/csp2.80.
Genersch, E., W. von der Ohe, H. Kaatz, A. Schroeder, C. Otten, R. Büchler, S. Berg, W. Ritter, W. Mühlen, S. Gisder, M. Meixner, G. Liebig, P. Rosenkranz, 2010: The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies. Apidologie 41, 332–352, DOI: 10.1051/apido/2010014.
Gérard, M., M. Vanderplanck, T. Wood, D. Michez, 2020: Global warming and plant-pollinator mismatches. Emerging topics in life sciences ETLS20190139, DOI: 10.1042/ETLS2019019.
Graystock, P., D. Goulson, W.O.H. Hughes, 2015: Parasites in bloom: flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proceedings of the Royal Society B: Biological Sciences 282 (1813), 20151371, DOI: 10.1098/rspb.2015.1371.
Hallmann, C.A., M. Sorg, E. Jongejans, H. Siepel, N. Hofland, H. Schwan, W. Stenmans, A. Müller, H. Sumser, T. Hörren, D. Goulson, H. De Kroon, 2017: More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLOS One, 12 (10), e0185809, DOI: 10.1371/journal.pone.0185809.
Hallmann, C.A., T. Zeegers, R. Van Klink, R. Vermeulen, P. Van Wielink, H. Spijkers, J. Van Deijk, W. Van Steenis, E. Jongejans, 2020: Declining abundance of beetles, moths and caddisflies in the Netherlands. Insect Conservation and Diversity 13 (2), 127–138,, DOI: 10.1111/icad.12377.
He, X.J., L.Q. Tian, X.B. Wu, Z.J. Zeng, 2016: RFID monitoring indicates honeybees work harder before a rainy day. Insect Science 23, 157–159, DOI: 10.1111/1744-7917.12298.
Hennessy, G., C. Harris, C. Eaton, P. Wright, E. Jackson, D. Goulson, F.F.L.W. Ratnieks, 2020: Gone with the wind: effects of wind on honey bee visit rate and foraging behavior. Animal Behaviour 161, 23e31, DOI: 10.1016/j.anbehav.2019.12.018.
IPBES, 2019: Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Global Assessment Report on Biodiversity and Ecosystem Services. IPBES Secretariat, Paris, France.
Jacques, A., M. Laurent, EPILOBEE Consortium, M. Ribière-Chabert, M. Saussac, S. Bougeard, G.E. Budge, P. Hendrikx, M.P. Chauzat, 2017: A pan-European epidemiological study reveals honey bee colony survival depends on beekeeper education and disease control. PLOS ONE 12 (3), e0172591, DOI: 10.1371/journal.pone.0172591.
James, R.R., 2005: Temperature and chalkbrood development in the alfalfa leafcutting bee, Megachile rotundata. Apidologie 36 (1), 15 – 23, DOI: 10.1051/apido:2004065.
Kerr, J.T., A. Pindar, P. Galpern, L. Packer, S.G. Potts, S.M. Roberts, P. Rasmont, O. Schweiger, S.R. Colla, L.L. Richardson, D.L. Wagner, L.F. Gall, D.S. Sikes, A. Pantoja, 2015: Climate change impacts on bumblebees converge across continents. Science 349, 177–180, DOI: 10.1126/science.aaa7031.
Kevan, P.G., B.F. Viana, 2018: The global decline of pollination services. Biodiversity 4 (4) 3–8, DOI: 10.1080/14888386.2003.9712703.
Lister, B.C., A. Garcia, 2018: Climate-driven declines in arthropod: abundance restructure a rainforest food web. Proceedings of the National Academy of Sciences USA 115, E10397.
Mallinger, R.E., H.R. Gaines-Day, C. Gratton, 2017: Do managed bees have negative effects on wild bees?: A systematic review of the literature. PLOS ONE 12 (12), e0189268, DOI: 10.1371/journal.pone.0189268.
Matias, D.M.S., C. Borgemeister, H. von Wehrden, 2017: Thinking beyond Western commercial honeybee hives: towards improved conservation of honey bee diversity. Biodiversity and Conservation 26, 3499–3504, DOI: 10.1007/s10531-017-1404-y.
Moritz, R.F.A., S. Erler, 2016: Lost colonies found in a data mine: Global honey trade but not pests or pesticides as a major cause of regional honeybee colony declines. Agriculture Ecosystems and Environment 216, 44-50, DOI: 10.1016/j.agee.2015.09.027.
Natsopoulou, M.E., D.P. McMahon, V. Doublet, E. Frey, P. Rosenkranz, R.J. Paxton, 2017: The virulent, emerging genotype B of Deformed wing virus is closely linked to overwinter honeybee worker loss. Scientific Reports 7, 5242, DOI: 10.1038/s41598-017-05596-3.
Nürnberger, F., S. Härtel, I. Steffan-Dewenter, 2019: Seasonal timing in honey bee colonies: phenology shifts affect honey stores and varroa infestation levels. Oecologia 189, 1121–1131, DOI: 10.1007/s00442-019-04377-1.
Ogilvie, J.E., S.R. Griffin, Z.J. Gezon, B.D. Inouye, N. Underwood, D.W. Inouye, R.E. Irwin, 2017: Interannual bumble bee abundance is driven by indirect climate effects on floral resource phenology. Ecology Letters 20, 1507-1515, DOI: 10.1111/ele.12854.
Potts, S.G., S.P.M. Roberts, R. Dean, G. Marris, M.A. Brown, R. Jones, P. Neumann, J. Settele, 2010: Declines of managed honey bees and beekeepers in Europe. Journal of Apicultural Research 49: 15–22, DOI: 10.3896/IBRA.1.49.1.02.
Powney, G.D., C. Carvell, M. Edwards, R.K.A. Morris, H.E. Roy, B.A. Woodcock, N.J.B. Isaac, 2019: Widespread losses of pollinating insects in Britain. Nature Communications 10, 1018, DOI: 10.1038/s41467-019-08974-9.
Puerta, F., J.M. Flores, M. Bustos, F. Padilla, F. Campano, 1994: Chalkbrood development in honeybee brood under controlled conditions. Apidologie 25 (6), 540–546, DOI: 10.1051/apido:19940604.
Sánchez-Bayo, F., K.A.G. Wyckhuys, 2019: Worldwide decline of the entomofauna: A review of its drivers. Biological Conservation 232, 8-27, DOI: 10.1016/j.biocon.2019.01.020.
Scaven, V.L., N.E. Rafferty, 2013: Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions. Current zoology 59 (3), 418–426, DOI: 10.1093/czoolo/59.3.418.
Schroeder, D.C., S.J. Martin, 2012: Deformed wing virus - The main suspect in unexplained honeybee deaths worldwide. Virulence 3 (7), 589-591, DOI: 10.4161/viru.22219.
Schulte, L.A., J. Niemi, M.J. Helmers, M. Liebman, J.G. Arbuckle, D.E. James, R.K. Kolka, M.E. O'Neal, M.D. Tomer, J.C. Tyndall, H. Asbjornsen, P. Drobney, J. Neal, G. van Ryswyk, C. Witte, 2017: Prairie strips improve biodiversity and the delivery of multiple ecosystem services from corn–soybean croplands. Proceedings of the National Academy of Sciences USA 114 (42),11247-11252, DOI: 10.1073/pnas.1620229114.
Seibold, S., M.M. Gossner, N.K. Simons, N. Blüthgen, J. Müller, D. Ambarlı, C. Ammer, J. Bauhus, M. Fischer, J.C. Habel, K.E. Linsenmair, T. Nauss, C. Penone, D. Prati, P. Schall, E.D. Schulze, J. Vogt, S. Wöllauer, W.W. Weisser, 2019: Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 574, 671-674, DOI: 10.1038/s41586-019-1684-3.
Soroye, P., T. Newbold, J. Kerr, 2020: Climate change contributes to widespread declines among bumble bees across continents. Science 367, 685–688, DOI: 10.1126/science.aax8591.
Steffan-Dewenter, I., T. Tscharntke, 2000: Resource overlap and possible competition between honey bees and wild bees in central Europe. Oecologia 122, 288–296, DOI: 10.1007/s004420050034.
Switanek, M., K. Crailsheim, H. Truhetz, R. Brodschneider, 2017: Modelling seasonal effects of temperature and precipitation on honey bee winter mortality in a temperate climate. Science of the Total Environment 579, 1581-1587, DOI: 10.1016/j.scitotenv.2016.11.178.
Tedesco, P.A., R. Bigorne, A.E. Bogan, X. Giam, L.C. Jézéque, B. Hugueny, 2014: Estimating how many undescribed species have gone extinct. Conservation Biology 28, 1360-1370, DOI: 10.1111/cobi.12285.
Theodorou, P., R. Radzevičiūtė, B. Kahnt, A. Soro, I. Grosse, R.J. Paxton, 2018: Genome-wide single nucleotide polymorphism scan suggests adaptation to urbanization in an important pollinator, the red-tailed bumblebee (Bombus lapidarius L.), Proceedings of the Royal Society B. Biological Sciences 285, 20172806, DOI: 10.1098/rspb.2017.2806.
Theodorou, P., R. Radzevičiūtė, G. Lentendu, B. Kahnt, M. Husemann, C. Bleidorn, J. Settele, O. Schweiger, I. Grosse, T. Wubet, T.E. Murray, R.J. Paxton, 2020: Urban areas as hotspots for bees and pollination but not a panacea for all insects. Nature Communications 11, 576, DOI: 10.1038/s41467-020-14496-6.
Thomson, J.D., 2001: Using pollination deficits to infer pollinator declines: Can theory guide us? Conservation Ecology 5 (1), 6.
Torchio, P.F., 1992: Effects of Spore Dosage and Temperature on Pathogenic Expressions of Chalkbrood Syndrome Caused by Ascosphaera torchioi within Larvae of Osmia lignaria propinqua (Hymenoptera: Megachilidae). Environmental Entomology 21 (5), 1086–1091, DOI: 10.1093/ee/21.5.1086.
von Büren, R.S., B. Oehen, N.J. Kuhn, S. Erler, 2019: High-resolution maps of Swiss apiaries and their applicability to study spatial distribution of bacterial honey bee brood diseases. PeerJ 7, e6393, DOI: 10.7717/peerj.6393.
Wagner, D.L., 2020: Insect Declines in the Anthropocene. Annual Review of Entomology 65 (1), 457-480, DOI: 10.1146/annurev-ento-011019-025151.
Whitfield, C.W., S.K. Behura, S.H. Berlocher, A.G. Clark, J.S. Johnston, W.S. Sheppard, D.R. Smith, A.V. Suarez, D. Weaver, N.D. Tsutsui, 2006: Thrice out of Africa: ancient and recent expansions of the honey bee, Apis mellifera. Science 314 (5799), 642–645, DOI: 10.1126/science.1132772.
Woodcock, B.A., J.M. Bullock, R.F. Shore, M.S. Heard, M.G. Pereira, J. Redhead, L. Ridding, H. Dean, D. Sleep, P. Henrys, J. Peyton, S. Hulmes, L. Hulmes, M. Sárospataki, C. Saure, M. Edwards, E. Genersch, S. Knäbe, R.F. Pywel, 2017: Country-specific effects of neonicotinoid pesticides on honey bees and wild bees. Science 356 (6345), 1393-1395, DOI: 10.1126/science.aaa1190.

 Suchen
Suchen